
Uvod
Povišen salinitet tla je rezultat prirodne akumulacije NaCl u tlu, ali može nastati i kao posljedica djelovanja čovjeka na okoliš kroz čitav niz agronomskih djelatnosti, primjerice navodnjavanjem, intenzivnom gnojidbom, krčenjem šuma ili pretjeranom ispašom („ overgrazing“) (Stavi i sur., 2021). Salinizacija narušava strukturu zemljišta, smanjuje aeraciju tla te ograničava opskrbu biljke vodom što u konačnici dovodi do pada prinosa (Dixit, 2022). Prema izvješću Agencije o stanju okoliša (AZO) za vremenski period od 2005-2008. godine, u Republici Hrvatskoj najveći udio zaslanjenih tala zabilježen je u području donje Neretve čija je prosječna električna provodljivost tla (EC) vezana za povećanje koncentracije Na+ i Cl- u 2007. godini bila za 64 % veća u odnosu na prethodnu godinu (AZO, 2012). S druge strane, u istom izvješću navodi se kako je pojava alkalizacije uglavnom vezana za područje Slavonije i Baranje, a zauzima 410 ha površine. Alkalna (halomorfna) tla nastaju kao posljedica vezivanja više od 15 % natrijevih iona za adsorpcijski kompleks tla, što povećava retencijski kapacitet tla i ograničava unos vode u korijen (Vidaček i sur., 2004).
Solni stres je rezultat taloženja NaCl i Na2SO4 te ostalih neutralnih soli u tlima što dovodi do povećanja koncentracije Na+ u biljnim stanicama te pojave osmotskog i ionskog stresa. Osmotski stres posljedica je unosa Na+ i Cl- što dovodi do pada vodnog potencijala tla, a visoke koncentracije Na+ izvan stanice reduciraju osmotski potencijal što dovodi do dehidracije stanica, pada turgora i promjene staničnog volumena (Sairam i Tyagi, 2004). Ova pojava je energetski zahtjevna čime se usporava rast i razvoj te mijenja metabolička aktivnost biljke (Fricke i Peters, 2002). Za razliku od osmotskog stresa, učinci ionskog stresa mogu značajno oštetiti funkciju i integritet stanice. Povišena koncentracija Na+ iona u stanici izaziva ionski stres, ometa unos hranjiva primjerice K+, PO43- i Ca2+ dok apsorpcija Cl- inhibira unos NO3- i H2PO4- (Munns i Tester, 2008; Lin i sur., 2018). Tolerantnost neke vrste na stres izazvan NaCl ne znači tolerantnost na stres izazvan Na2SO4. Brojni autori naglašavaju veću osjetljivost na Na2SO4 u odnosu na NaCl pri istim izotoničnim uvjetima u biljkama soje (Boursier i Läuchli, 1990) i pšenice (Datta i sur., 1995).
Stres izazvan alkalnim solima (NaHCO3 i Na2CO3) uzrokuje slične promjene kao i stres izazvan neutralnim solima (NaCl i Na2SO4) osim što dodatno povećava pH vrijednost tla (Wang i sur., 2022). Povećanje pH vrijednosti u tlu uzrokuje taloženje dvovalentnih iona (Fe2+, Mg2+ i Ca2+) koji ometaju unos aniona i ostalih hranjivih tvari čime narušavaju hranidbeni status biljke (Chen i sur., 2017). Povećan udio HCO3- i SO42-u tlima rezultat je evaporacije vode ili može biti rezultat prirodnog ispiranja stijena (Stavi i sur., 2021). U bermudske trave ( Cynodon dactylon (L.) Pers.) slaba alkalizacija uzrokovala je sporiju degradaciju šećera i smanjenu aktivnost metabolizma dušika, dok su umjeren te jaki intenzitet alkalizacije potakli veću akumulaciju osmotski aktivnih šećera, veću produkciju ROS čestica i veća oštećenja membrana (Ye i sur., 2021). Salinitet i alkalitet negativno utječu na svaku razvojnu fazu biljke no faza klijanja je najosjetljivija jer klica dolazi u direktni kontakt s vodenom otopinom soli u tlu (Hu i sur., 2018). Stoga se praćenje germinacije smatra relativno pouzdanim screening testom u istraživanju tolerantnosti saliniteta i alkaliteta neke vrste, odnosno genotipa. Salinitet i alkalitet usporavaju klijanje i produžuju vrijeme imbibicije sjemena (Munns, 2002). Povećane koncentracije Na+ i Cl- mijenjaju ravnotežu hormona uključenih u regulaciju stanja dormantnosti i klijanja. Tako, salinitet povećava endogenu koncentraciju apscizinske kiseline (ABA), hormona dormantnosti, a istovremeno inhibira sintezu hormona giberelina (GAs), stimulatora klijanja (Lee i Luan, 2012).
Industrijska konoplja ( Cannabis sativa L.) je jednogodišnja, dvodomna biljka koja ima široku primjenu u brojnim industrijama. Konoplja je kultura koja posjeduje tolerantnost na salinitet, ali postoji velika genetička varijabilnost između kultivara (Dixit, 2022). Stoga je cilj ovog rada bio istražiti učinak neutralnih (NaCl, Na2SO4) i alkalnih (NaHCO3, Na2CO3) soli primijenjenih u jednakoj koncentraciji (100 mM) na klijanje i rast korijena dva genotipa industrijske konoplje (Santhica 70 i Futura 83) tijekom četiri dana rasta.
Materijal i metode
Eksperimentalni postav
U istraživanju su korištena dva kultivara industrijske konoplje Santhica 70 i Futura 83 (Organica Vita, Čađavica, Republika Hrvatska). Za provjeru klijavosti, sjeme oba kultivara postavljeno je u plastične prozirne posude s poklopcem (20 cm x 15 cm x 3 cm) te dvoslojnim filter papirom na dnu posude. Prije postavljanja sjemena, u kontrolne posudice dodana je dH2O, posude sa otopinama NaCl i Na2SO4 su predstavljale solni stres, dok je alkalni stres izazvan dodatkom otopina NaHCO3 i Na2CO3. Koncentracija svih otopina iznosila je 100 mM, a volumen dodane tekućine u svakoj posudi iznosio je 10 mL. Na filter papir je potom u pravilnim razmacima postavljeno 30 sjemenki odabranih kultivara konoplje tako da budu podjednake veličine i bez mehaničkog oštećenja. U posudama sa solnim i alkalnim otopinama filter papir je mijenjan svaka dva dana kako bi očuvali konstantnu koncentraciju soli na podlozi dok su kontrolne posude svaka dva dana bile zalijevane s dH2O. Nakon naklijavanja sjeme je prva dva dana bilo u tami, a nakon toga je izloženo 12/12 satnoj umjetnoj rasvjeti fluorescentnih svjetiljki (80 μmol fotona m-2 s-1) u sobi za uzgoj pri 23 ± 2 °C tijekom četiri dana trajanja eksperimenta.
Praćenje klijavosti i rasta korijena
Klijavost sjemena evidentirana je 2. i 4. dana uzgoja , a kao kriterij klijavosti uzeta je pojavnost korjenčića. Postotak klijavosti (G) određen je koristeći se izračunom (Thabet i sur., 2018) (1);
G (%) = (1)
Gdje, n predstavlja broj proklijalih sjemenki, a N ukupan broj sjemenki u jednoj posudi. Germinacijski indeks tolerantnosti na sol (STI_G, germination salt tolerance index) izračunat je za 2. i 4. dan klijanja na temelju 90 sjemenki prema jednadžbi (Wu i sur., 2019) (2);
STI = (2)
Duljina korijena mjerenja je 4. dana rasta pomoću ravnala na 10 reprezentativnih uzoraka po posudi i tretmanu. Svaki tretman i kontrola rađeni su u tri ponavljanja za svaki kultivar pojedinačno.
Obrada podataka
Rezultati svih mjerenja izraženi su kao srednje vrijednosti od najmanje tri do najviše 10 repeticija po genotipu i tretmanu ovisno o analizi uz prikaz standardne devijacije (± S.D.). Za statističku obradu podataka korištena je jednofaktorijalna analiza varijance (One-Way ANOVA). Statistički značajne razlike između tretmana i kultivara provjerene su uz pomoć Post hoc LSD (engl. Least Significant Difference) testa (Statistica 14.0.0.15) na razini značajnosti P≤0,05.
Rezultati i rasprava
Učinak saliniteta i alkaliteta na klijavost
Neutralni i alkalni solni stres uzrokovali su inhibiciju klijavost u odnosu na kontrolu u oba genotipa u obje vremenske točke (Slika 1.). Izuzetak je bio kultivar Futura 83 kod kojeg nakon 4. dana uzgoja (G4) nije bilo statistički značajne razlike između sjemena koje je raslo na kontroli i sjemena na tretmanu NaHCO3 iz čega slijedi kako je alkalni stres izazvan otopinom NaHCO3 potakao bržu germinaciju. Slično je uočeno i kod soje kod koje je povećana pH vrijednost uzrokovana alkalnim stresom (NaHCO3) potaknula brže otvaranje sjemene lupine i ubrzala germinaciju (De Lacerda i sur., 2003). Nakon 2. dana uzgoja, kod kultivara Santhica 70 uočen je značajni porast klijavosti na NaCl tretmanu u odnosu na ostale soli, dok je sjeme Future 83 pokazalo podjednako dobru klijavost na NaCl i NaHCO3 tretmanima. Neki autori naglašavaju kako toleranciji saliniteta u fazi klijanja doprinosi anatomija sjemene lupine, struktura i veličina sjemenke (Demir i Mavi, 2008; Hu i sur., 2018). Na temelju ovoga može se pretpostaviti da je različita klijavost ispitivanih kultivara konoplje na solnom i alkalnom stresu posljedica različite topografije i strukture sjemena što zahtjeva dodatna istraživanja. Nadalje, primjena alkalnih otopina (100 mM Na2CO3 i 100 mM NaHCO3) uzrokovala je jača oštećenja staničnih membrana kod pšenice nego stres izazvan neutralnim otopinama (100 mM Na2SO4 i 100 mM NaCl) (Liu i sur., 2022). Najveća inhibicija klijavosti u ranoj fazi (G2) uočena je na tretmanu Na2SO4 u odnosu na kontrolu i ostale otopine soli. Tako je, klijavost bila manja za 97,6 % (Santhica 70) i 90,6 % (Futura 83) u odnosu na kontrolu, te 75 % (Santhica 70) i 61,5 % manja (Futura 83) od sjemena koje je raslo na Na2CO3. Santhica 70 pokazala je i značajno smanjene vrijednosti STI_G2 u odnosu na Futuru 83 (Slike 2 I. i 2 III.). Nadalje, nakon 4. dana rasta, samo je kod Future 83 uočen različit učinak neutralih soli na klijanje; sjeme uzgajano na otopini NaCl bolje je klijalo i pokazalo veći STI_G4 nego na tretmanu Na2SO4 (Slika 2 IV.). Ovo je u skladu s istraživanjem provedenom na konoplji kod koje je primjena 150 mM otopine Na2SO4 imala jači negativni učinak na postotak klijavosti nego 150 mM otopina NaCl (Hu i sur., 2018). U uvjetima stresa biljke pojačano sintetiziraju polifenole, koji pripadaju neenzimatskom antioksidacijskom sustavu obrane. Tako je kod vrste P. strombulifera uočena veća koncentracija polifenola u tretmanu otopinom Na2SO4 u odnosu na otopinu NaCl (Reginato i sur., 2014). Stoga, jača inhibicija klijanja i rasta pri Na2SO4 kod ispitivanih kultivara konoplje može biti posljedica i pojačane akumulacije polifenolnih spojeva što se treba dodatno potvrditi. Uspoređujući postotak klijavosti između genotipova, vidljivo je da je Futura 83 u ranijoj vremenskoj točki (G2) imala značajno veće vrijednosti na kontroli i svim solnim tretmanima od kultivara Santhica 70 (Slika 1.). Kasnije (G4), taj trend je uočen samo na tretmanima NaCl i NaHCO3 što potvrđuju i značajno veće vrijednosti STI_G2 i STI_G4 kod kultivara Futura 83 (Slike 2 I. i 2 III.). Iz ovoga slijedi kako Futura 83 pokazuje veću tolerantnost saliniteta uzrokovanog NaCl i alkaliteta izazvanog NaHCO3, od kultivara Santhica 70. Slično je uočeno i kod osjetljivog kultivara industrijske konoplje (var. Bamahuoma) gdje je solni stres (100-300 mM NaCl) uzrokovao inhibiciju klijavosti u odnosu na tolerantan genotip (YM5) (Hu i sur., 2018).
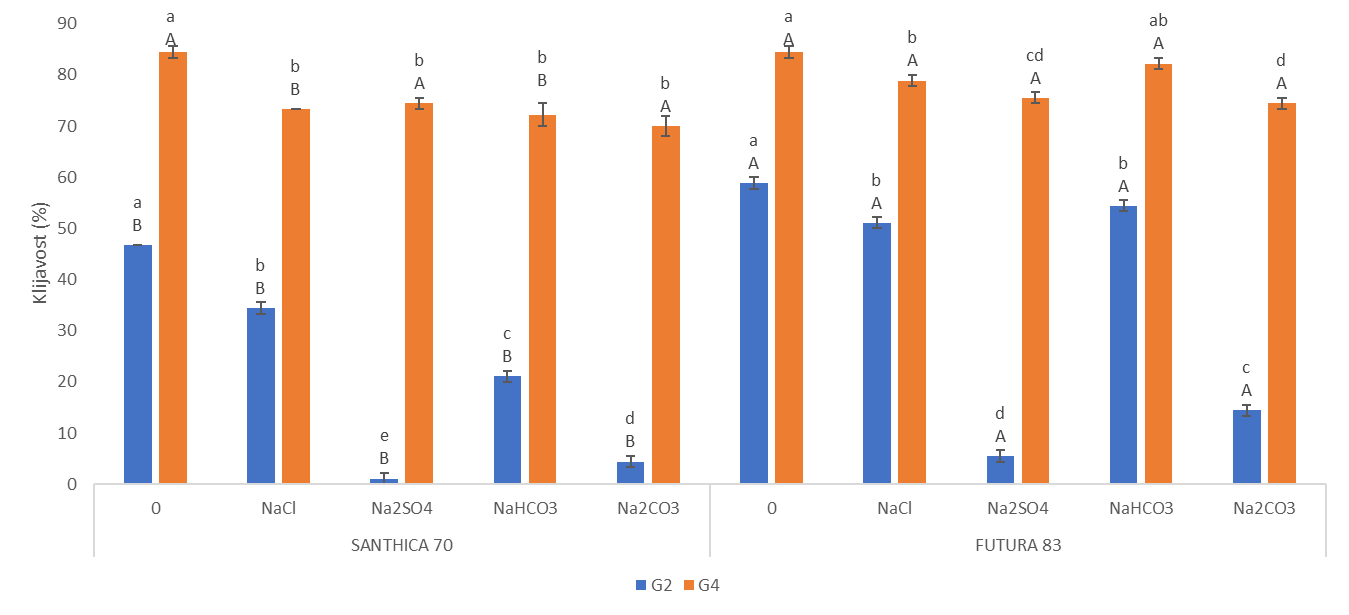
Slika 1. Učinak saliniteta (NaCl i Na2SO4) i alkaliteta (NaHCO3 i Na2CO3) na klijavost dva kultivara industrijske konoplje (Santhica 70 i Futura 83) na kontroli i tretmanu 100 mM nakon 2. dana (G2) i 4. (G4) dana uzgoja. Vrijednosti su prikazane kao srednja vrijednost 3 replike ± S.D. Velika slova iznad stupaca predstavljaju značajne razlike između kultivara na istom tretmanu, a mala slova označuju razlike između tretmana unutar jednog kultivara za jednu vremensku točku (P≤ 0,05, LDS test).
Figure 1. Effect of salinity (NaCl i Na2SO4) and alkalinity (NaHCO3 i Na2CO3) on germination in two industral hemp cultivars (Santhica 70 and Futura 83) under control and 100 mM treatment after 2. day (G2) and 4. day (G4) of growth. Values present means of 3 repetition ± S.D. Upper case letters above bars indicate significant differences between cultivars under same treatment whilst lower case letters show differences between treatments within each cultivars for one separate time point (P≤ 0,05, LDS test).
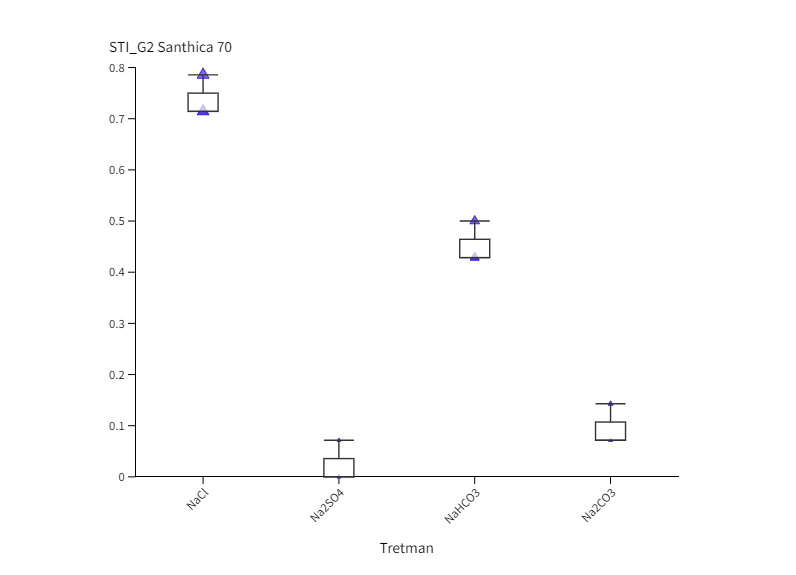
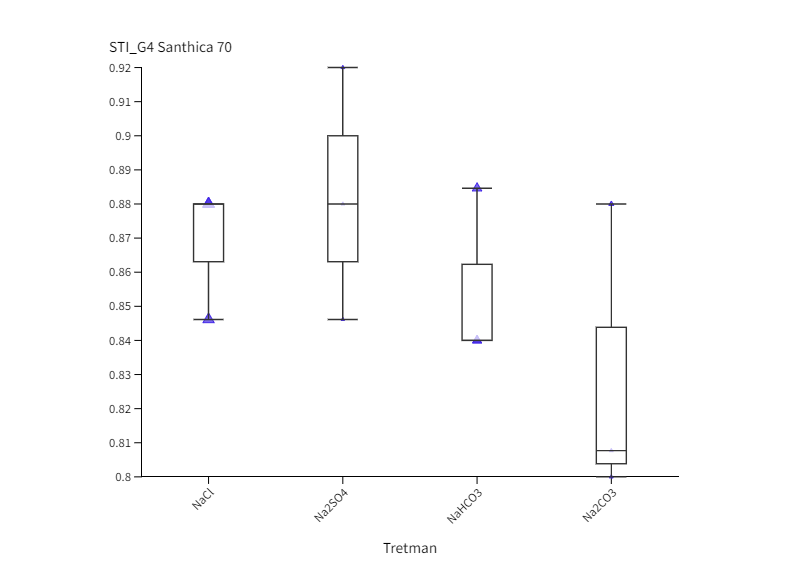
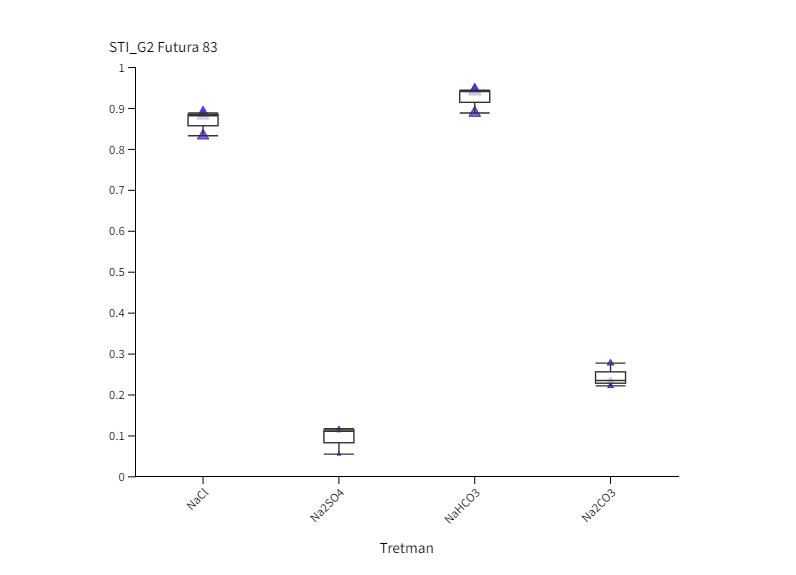
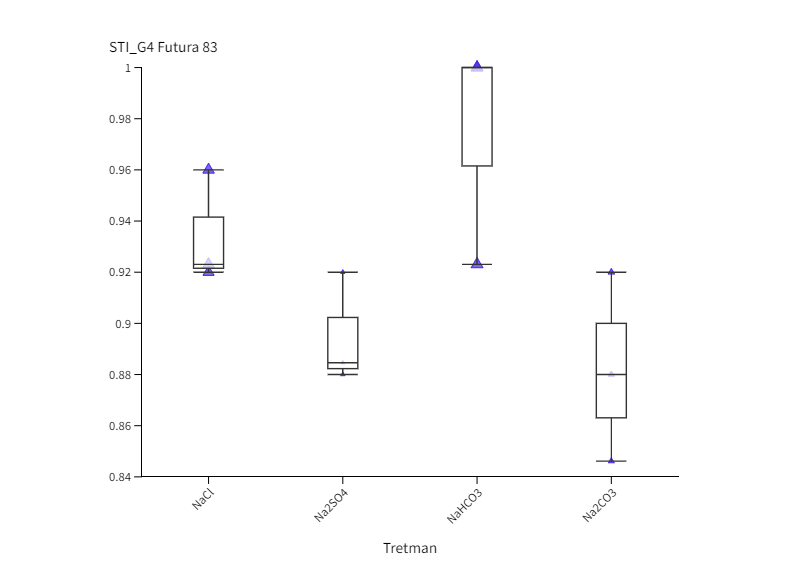
Slika 2. Box-plot projekcija učinka saliniteta (NaCl i Na2SO4) i alkaliteta (NaHCO3 i Na2CO3) na germinacijski indeks tolerancije saliniteta (STI_G) kod kultivara Santhica 70 ( I., II.) i Futura 83 ( III., IV.) nakon 2. dana (STI_G2) i 4. dana (STI_G4) uzgoja. Velika slova iznad stupaca predstavljaju značajne razlike između kultivara na istom tretmanu, a mala slova označuju razlike između tretmana unutar jednog kultivara za jednu vremensku točku (P≤ 0,05, LDS test).
Figure 2. Box-plot projection of salinity (NaCl and Na2SO4) and alkalinity (NaHCO3 and Na2CO3) effect on germination in two industrial hemp cultivars (Santhica 70 and Futura 83) under control and 100 mM treatment after 2. (G2) and 4. day (G4) of growth. Upper case letters above bars indicate significant differences between cultivars under same treatment whilst lower case letters show differences between treatments within each cultivar for one separate time point (P≤ 0,05, LDS test).
Učinak saliniteta i alkaliteta na rast korijena
Svi solni tretmani imali su negativan učinak na rast korijena u odnosu na biljke uzgojene na kontrolnom tretmanu (Slika 3.). Duljina korijena u oba kultivara bila je najkraća primjenom otopine Na2CO3, a najduža u biljaka koje su rasle pri otopini NaCl (Slika 4.). Slično kao i kod germinacije, otopina NaCl imala je najslabiji inhibitorni učinak na rast kod oba kultivara. Uspoređujući učinak neutralnih soli na rast korijena, vidljivo je da je korijen uzgojen na Na2SO4 tretmanu bio značajno kraći nego na otopini NaCl iz čega slijedi da odgovor na salinitet ovisi o anionskom sastavu soli. Sulfati predstavljaju glavni izvor sumpora koji je ključan makroelement u rastu i razvoju biljke. Redukcijom sulfata u procesu asimilacije u plastidima nastaje aminokiselina cistein koja služi kao skladište organskog sumpora (De Kok i sur., 2005). U nizu konverzijskih koraka sulfat se uz pomoć enzima 5'-fosfosulfat reduktaze (APR) prevodi do otrovnog intermedijera sulfita, potom do sulfida uz pomoć sulfitne reduktaze (SiR) i konačno do cisteina uz pomoć 0-acetilserin(tiol) liaze (Yarmolinsky i sur., 2013). Iz ovoga se može pretpostaviti da su povećane koncentracije sulfatnog aniona uzrokovale inhibiciju enzima APR čime je došlo do nagomilavanja štetnog SO32-. Suprotno od učinka soli na germinaciju, izlaganje korijena otopini Na2CO3 uzrokovalo je značajnu redukciju rasta u odnosu na sve solne tretmane kod oba genotipa što može biti posljedica narušavanja selektivnosti membrane stanica korijena uslijed porasta pH vrijednosti izazvanog alkalnim stresom. Naime, niska kiselost ometa selektivnu K+/Na+ apsorbciju u korijenu što narušava unutarstaničnu K+/Na+ koncentraciju (Chen i sur., 2012). Jedan od adaptivnih mehanizama tolerantnosti alkaliteta je i povećana aktivnost H+-ATP-aza čijim se radom zakiseljuje apoplast i inducira rast korijena (Sánchez-Rodríguez i sur., 2010; Chen i sur., 2017). Zakiseljavanje apoplasta inducira sintezu hormona auksina koji potiče elongaciju, odnosno rast korijena u dužinu (Chen i sur., 2017). Stoga je moguće da je inhibicija rasta korijena uvjetima povišenog alkaliteta posljedica smanjene aktivnosti protonskih pumpi (H+-ATP-aza). Slično rezultatima dobivenim ovim istraživanjem, visina biljke, suha tvar i duljina korijena kod konoplje imale su niže vrijednosti pri tretmanu s Na2CO3 u odnosu na NaCl (Hu i sur., 2019). Ipak, navedeni genotipovi pokazali su jednaku inhibiciju rasta pri ekstremno visokoj koncentraciji NaCl (300 mM), dok je izlaganje 300 mM Na2CO3 imalo letalan učinak na rast. Iz ovoga se može zaključiti kako tolerantnost alkaliteta i saliniteta kod konoplje ovisi o vrsti soli i intenzitetu stresa. Jača oštećenja strukture korijenovog sustava nakon izlaganja smjesi alkalnih soli (NaHCO3 i Na2CO3) u odnosu na smjesu neutralnih soli (NaCl i Na2SO4) uočena su i u suncokreta ( Helianthus annuus L.) (Liu i sur. 2010). Genotipske razlike u duljini korijena uočene su na kontroli gdje je Santhica 70 razvila dulji korijen od Future 83 dok je na tretmanu NaCl korijen genotipa Futura 83 bio značajno duži (Slika 3.) iz čega se može zaključiti da ovaj kultivar uspješnije tolerira solni stres izazvan NaCl. Razlike u toleriranju saliniteta uočene su i između dva kultivara konoplje (BM i YM5), gdje je kultivar BM pokazao manju inhibiciju rasta pri izlaganju otopinama 100 mM Na2CO3 i 100 mM NaCl te jaču indukciju stresnih proteina (Hu i sur., 2018; Hu i sur., 2019).
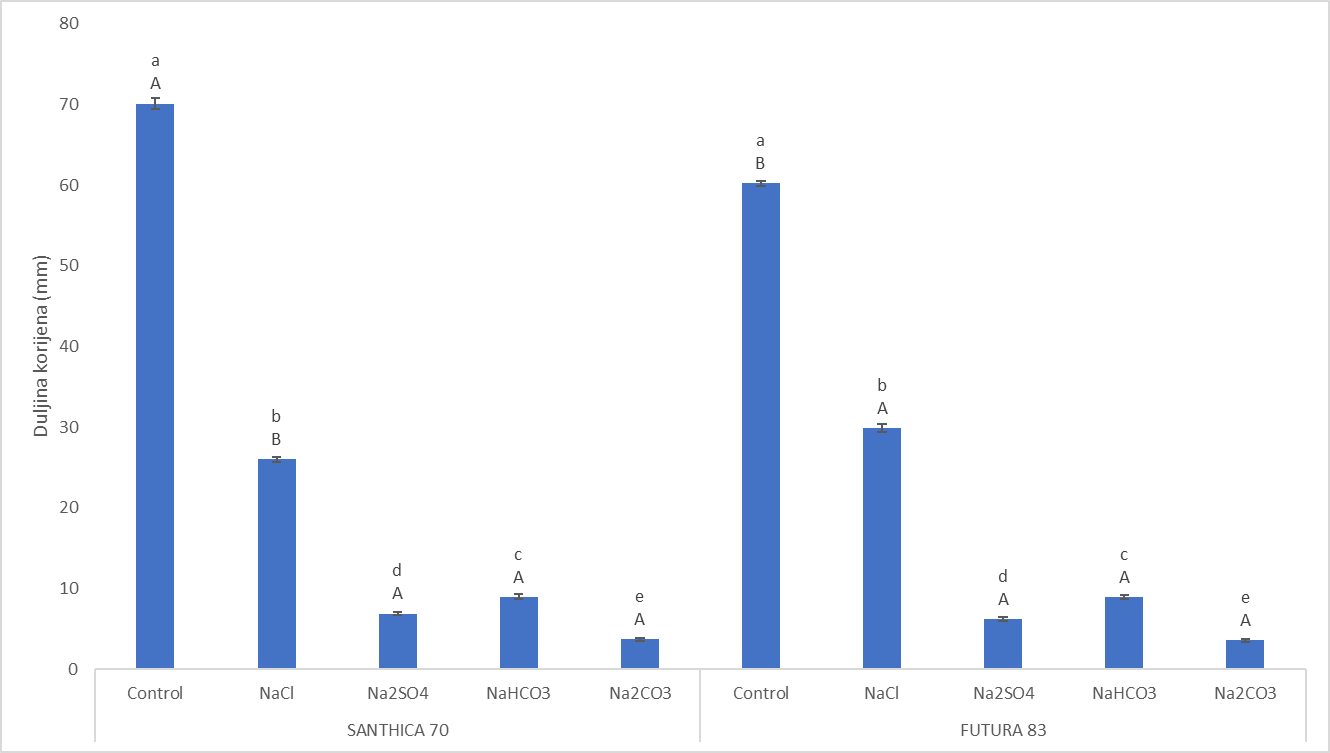
Slika 3. Učinak saliniteta (NaCl i Na2SO4) i alkaliteta (NaHCO3 i Na2CO3) na kontroli i tretmanu 100 mM na rast korijena dva kultivara industrijske konoplje (Santhica 70 i Futura 83) te kod kontrolnog sjemena nakon 4. dana uzgoja. Vrijednosti su prikazane kao srednja vrijednost 10 replika ± S.D. Velika slova iznad stupaca predstavljaju značajne razlike između kultivara na istom tretmanu, a mala slova označuju razlike između tretmana unutar jednog kultivara (P≤0,05, LSD test).
Figure 3. Effect of salinity (NaCl i Na2SO4) and alkalinity (NaHCO3 i Na2CO3) on root growth in two industral hemp cultivars (Santhica 70 and Futura 83) under control and 100 mM treatment after 4. day (G4). Values present means of 10 repetitions ± S.D. Upper case letters above bars indicate significant differences between cultivars under same treatment whilst lower case letters show differences between treatments within each cultivar for one separate time point (P≤0,05, LDS test).
NaCl
Na2SO4
NaHCO3
Na2CO3
Slika 4. Rast korijena na neutralnom (NaCl, Na2SO4) i alkalnom (NaHCO3 i Na2CO3) stresu kod kultivara Futura 83 pri koncentraciji 100 mM nakon 4 dana.
Figure 4. Root growth under neutral (NaCl, Na2SO4) and alkaline (NaHCO3 i Na2CO3) stress in cultivar Futura 83 under 100 mM after 4 days.
Zaključak
Salinitet (NaCl i Na2SO4) i alkalitet (Na2CO3 i NaHCO3) uzrokovali su značajnu redukciju postotka klijavosti i inhibiciju rasta korijena u odnosu na kontrolne biljke u oba kultivara konoplje. Nakon 2. dana uzgoja, solni stres izazvan otopinom Na2SO4 pokazao je najizraženiji inhibitorni učinak na klijavost te najniže vrijednosti STI za oba kultivara, dok su štetne posljedice solnog stresa potaknutog otopinom NaCl, bile najslabije vidljive. Za razliku od germinacije, korijen oba kultivara pokazao je najjaču redukciju rasta nakon izlaganja otopini Na2CO3, dok je korijen uzgojen na otopini NaCl rastao najbolje. Iz ovog slijedi da učinak različitih vrsta soli ovisi o biljnom organu. Tijekom vremena (4. dan uzgoja) kultivar Futura 83 imao je veći postotak klijavosti od kultivara Santhica 70 na tretmanima NaCl i NaHCO3, veće vrijednosti STI_G2 i STI_G4 te dulji korijen na tretmanu izazvanom otopinom NaCl, iz čega slijedi da Futura 83 pokazuje veću tolerantnost alkaliteta i saliniteta. Dobivena istraživanja mogu poslužiti u probiru tolerantnih genotipova konoplje na stres izazvan alkalitetom i salinitetom.