
Introduction
It is a broadly acknowledged theory that the fields around the large Tethys Sea (which covered part of the present Mediterranean zone during the Tertiary) became a center of speciation for many plant families after the Alpine uplift and Oligocene climatic changes during the Pleistocene and Holocene. Only several legume tribes such as Hedysareae, Genisteae, Viciae, Loteae, and Trifoliae, were able to endure the severe climatic conditions, surviving and multiplying under extreme pressure (summer drought, winter frost). The Fabaceae family of flowering plants, which includes roughly 19,500 species divided into 751 genera, is regarded as the third largest flowering plant family. This enormous family has demonstrated a high degree of species diversity and evolutionary success in a variety of global ecosystems, including stony grasslands, deserts, seashores, alpine and arctic meadows (Choi and Ohashi 2003, Legume Phylogeny Working Group 2023). The Mediterranean and Irano-Turanic phytogeographical regions have some specific genera such as Onobrychis, Hedysarum, Medicago, Astragalus, Anthyllis, Trigonella, Coronilla, Hippocrepis, Ononis, Genista, etc. (Lesins and Lesins 1979, Polhill 1981). Two areas are distinctly bipolar for this speciation: the eastern Mediterranean and Irano-Turanic zone (around the Aegean, Black, and Caspian seas), and the western Mediterranean or lbero-Maghrebi (around the Tyrrhenian and Alboran seas). Within the genus Hedysarum, circum-Mediterranean, mostly eastern Mediterranean, species are outstanding (among the 21 taxa recognized for Europe by Chrtková-Zertova, 1968, 13 are eastern Mediterranean). Among the 45 taxa recognized for the circum-Mediterranean areas by Greuter et al. (1989), 33 are present in the eastern Mediterranean.
In Türkiye alone, 22 taxa (one of which is suspect) of Hedysarum were recognized by Hedge (1979), and with the addition of the new taxa, the number of taxa in the genus will reach 24 (Bașköse et al. 2018, Aytaç et al. 2020, Hamzaoğlu and Koç 2020, Kandemir et al. 2023). Hedge (1979) separated Hedysarum into five sections ( Hedysarum, Obscura, Multicaulia, Subacaulia, and Crinifera) in the Flora of Türkiye. Of these, 11 are endemic to Türkiye, and the endemism ratio of the genus is 52.3%. The classification of sections of Hedysarum is also quite problematic. Until recently, it has included four sections: Hedysarum, Membranacea B. Fedtsch. (1902: 229), Stracheya (Benth.) B.H. Choi & H.Ohashi , and Multicaulia (Boiss.) B. Fedtsch. However, section Multicaulia consists of three subsections: Multicaulia B.H. Choi & H. Ohashi, Crinifera (Boiss.) B.H. Choi & H. Ohashi, and Subacaulia (Boiss.) B.H. Choi & H. Ohashi.
Characters such as the size, shape, and number of leaflets, indumentum, flower color, size, shape, and proportions of the standard, wings and keel, and shape and indumentum of the lomentum are extremely variable among the species of the Hedysarum genus (Hedge 1979). Hedysarum varium is the most common and polymorphic species of the genus in Türkiye. Hedge (1979) also noted that H. varium may be extended by hybridization or introgression with H. pestalozzae, H. syriacum, and H. nitidum. The differences between these four species are unsatisfactory, and further work is needed before their interrelationships and taxonomy can be placed on sounder foundations.
The palynomorphology of the many Fabaceae taxa has taxonomic significance (Erdtman 1969, Ohashi 1971, Perveen and Qaiser 1998, Pavlova and Manova 2000, Pınar et al. 2000, Avcı et al. 2013, Çeter et al. 2013, Bagheri et al. 2019). Three apertures, reticulate or suprareticulate ornamentation of exine, oblate-spheroidal or prolate pollen grains were given in those studies. Additionally, the micromorphological (pollen, fruit, and seed) characteristics can contribute to the discrimination of Hedysarum species, especially the pollen grains and muri size. However, there has been no taxonomical study conducted on the pollen structures of the subsection Multicaulia in Türkiye (Ohashi 1971, Polhill 1981, Faegri et al. 1989, Moore et al. 1991, Choi and Ohashi 1996, Civelek et al. 1999, Pavlova and Manova 2000, Sa et al. 2010, Ghanavati and Amirabadizadeh 2012, Dural and Citak 2015).
In order to enhance the pollen morphological research into the genus Hedysarum, the current study was conducted to describe and evaluate in detail, for the first time, the palynomorphological properties of seven taxa distributed in Türkiye using light microscopy (LM) and scanning electron microscopy (SEM). The objectives of this work were to: 1) identify and investigate the features of the pollen grains of species belonging to the Multicaulia subsection; and 2) use numerical analyses to clarify the systematic significance of the palynological traits.
Plant material
The specimens of the section Multicaulia were collected from various localities, as listed in Table 1, and stored in the herbarium of the Department of Biology of Selçuk University (KNYA). The taxonomical description of the species followed that of Hedge (1979).
Tab. 1. Localities of specimens of Hedysarum from the section Multicaulia collected from various localities in Türkiye and stored in KNYA herbarium.
Palynological analysis
Both LM and SEM were used to examine pollen grains of the genus Hedysarum. For palynological analysis, pollen samples of specimens were obtained from herbarium materials. To observe and calculate the characteristics of pollen grains, the Wodehouse technique was used (Wodehouse 1935). In this unique and easy technique, pollen grains were separated from the anthers and were stained with glycerin-jelly and covered with coverslips. A Leica DM 1000 light microscope with a Canon 450D camera (Ota City, Tokyo, Japan) and software from the Kameram 21 program (Argenit, Istanbul, Türkiye) was used to measure and observe fifty pollen grains per specimen. The equatorial diameter (E), polar axis (P), colpus length (Clg), colpus width (Clt), apocolpium (t), thickness of the exine and intine, and lumina width (LW) were examined and the P/E ratios were calculated. The mean, standard deviation (SD), minimum–maximum range, and mean values of these pollen characteristics for the taxa under study were given.
The unacetolyzed pollen grains were transferred directly onto aluminum stubs, covered with gold using a Cressington Auto 108 sputter coater (Cressington Scientific Instruments, Watford, Hertfordshire, UK) and photographed using a Zeiss Evo Ls10 scanning electron microscope (SEM) (Carl Zeiss NTS GmbH, Oberkochen, Germany) for the SEM analyses (Dural and Citak 2015).
The pollen terminologies of Punt et al. (2007), and Halbritter et al. (2018) were followed.
Statistical analysis
The determined qualitative and quantitative characters were scored for numerical analysis. Nine pollen characters were used to evaluate Hedysarum taxonomic relationships. These characters are equatorial and polar axes, exine, intine, colpus length, colpus width, apocolpium, sculpture and lumen width. Next, the determined qualitative and quantitative characters were turned into a data matrix. For the pollen characters of the seven taxa, the coefficients of correlation were detected and compiled using the clustering analysis method (unweighted pair group method with arithmetic mean (UPGMA), dissimilarity, standardized variables). Using nine characters and seven taxa, a primary matrix was established for the multivariate analysis. The Gower general coefficient similarity (Gower 1971), which can be utilized directly to a variety of character types (binary, qualitative, and quantitative characters), provided a basis for the clustering analysis. The UPGMA method was selected due to its widespread usage, congruence with the classification derived by traditional techniques, and visible accuracy in generating a reflection similarity matrix, as demonstrated by the co-phenetic correlation coefficient of Sokal and Rohlf (1962) and symmetrical hierarchical structure (Sokal and Rohlf 1962, McNeill 1979). The covariance matrix was created using non-standardized, untransformed, and centered data (Citak 2019). All calculations were performed via MVSP 3.22 software (Kovach Computing Services, Anglesey, Wales, Kovach 2013).
Pollen morphology
The pollen grains of the taxa of the subsection Multicaulia were determined as monad, radially symmetrical and isopolar. The shapes of the pollen grains were elliptic and not compressed at the poles in equatorial view or orbicular in polar view (Fig. 1, Fig. 2).
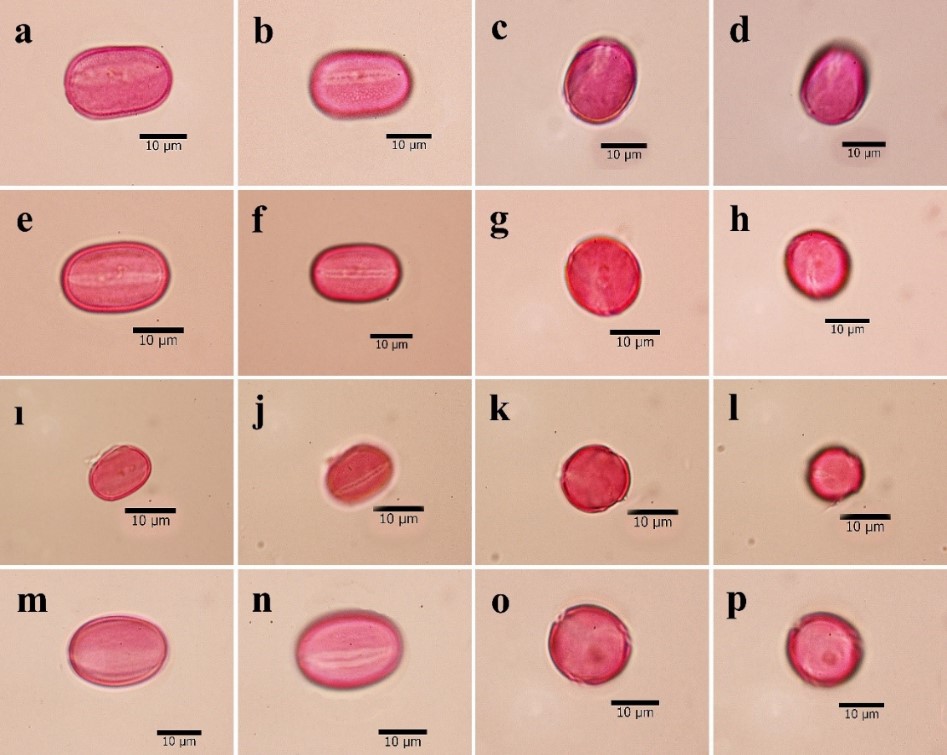
Fig. 1. Pollen microphotographs of species of the Hedysarum genus of Multicaulia: a-d. H. laxum, e-h. H. leucocladum, ı-l. H. huetii, m-p. H. nitidum.
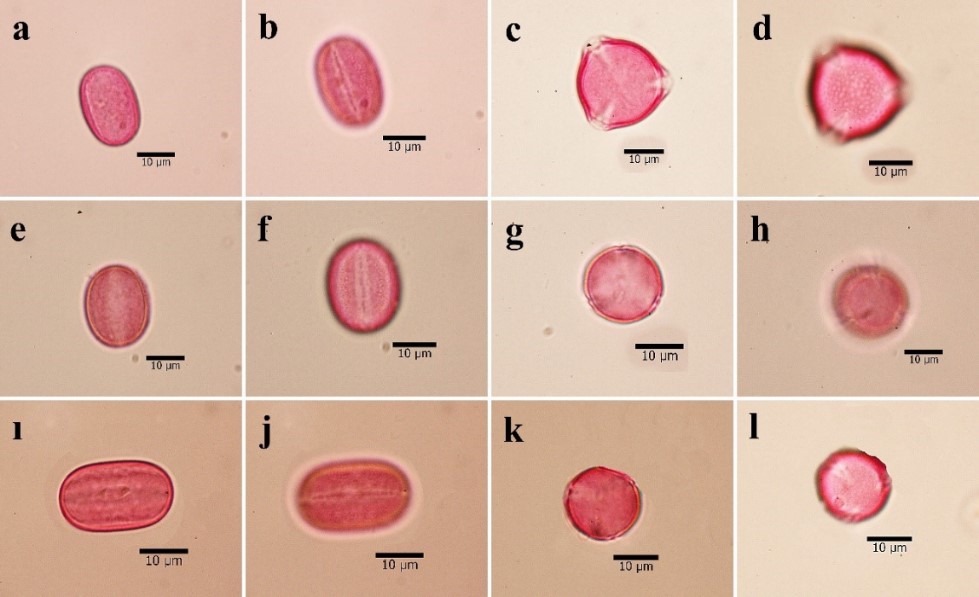
Fig. 2. Pollen microphotographs of species of Hedysarum genus of Multicaulia a-d. H. pestalozzae, e-h. H. varium subsp. varium ı-l. H. varium subsp. syriacum.
Quantitative characteristics of pollen grains from the plant samples are summarized in Table 2.
Tab. 2. The palynomorphological characters examined in Multicaulia subsection (values in µm, except for P/E). PS: pollen shape, P: prolate, SP: subprolate, E: exine, I: intine, Clg: colpus length, Clt: colpus width, t: apocolpium, Sc: sculpturing, MP: microreticulate-perforate, RP: reticulate-perforate, LW: lumina width. Fifty pollen grains per specimen were analyzed. * indicates endemic taxa and **indicates characters used in statistical analysis.
The general pollen shape in this study was determined as prolate; subprolate pollen shape was observed only in H. nitidum. The highest average pollen grain diameter on polar axis was observed in H. laxum (22.85 µm), whereas the lowest diameter was observed in H. huetii (20.53 µm). Similarly, the maximum pollen grain diameter on the equatorial axis was observed in H. nitidum (16.23 µm), while the minimum diameter was examined in H. laxum (12.68 µm). The size of the pollen grains was calculated based on the P/E ratio. The highest P/E value was observed in H. laxum (1.80), while the lowest value was observed in H. nitidum (1.33).
All of the studied members of the subsection Multicaulia had tricolpate apertures. The investigated species had different aperture sizes, with the maximum average colpi length observed in H. pestalozzae (19.84 µm), while the minimum was found in H. huetii (16.83 µm). In terms of the width, the maximum colpus width was found in H. nitidum (3.93 µm), while the minimum width was observed in H. leucocladum (2.97 µm). An operculum with psilate ornamentation on the colpus was determined in all pollen grains.
The exine ornamentation was the most distinguishing character of the species. In current study, all of the species had microreticulate-perforate exine sculpturing, except for H. varium subsp. varium, which had reticulate-perforate exine sculpturing (Fig. 3). The exine thickness ranged from a maximum of 1.38 µm in H. nitidum to a minimum of 0.96 µm in H. varium subsp. varium. The lumina width also varied among the species, and ranged from a maximum of 1.10 µm in H. varium subsp. varium to a minimum of 0.50 µm in H. pestalozzae. The sculpture of pollen grains was microreticulate-perforate or reticulate-perforate in equatorial view; however, only perforate ornamentation was observed in polar view. The walls of the muri were sinuous in the examined species.
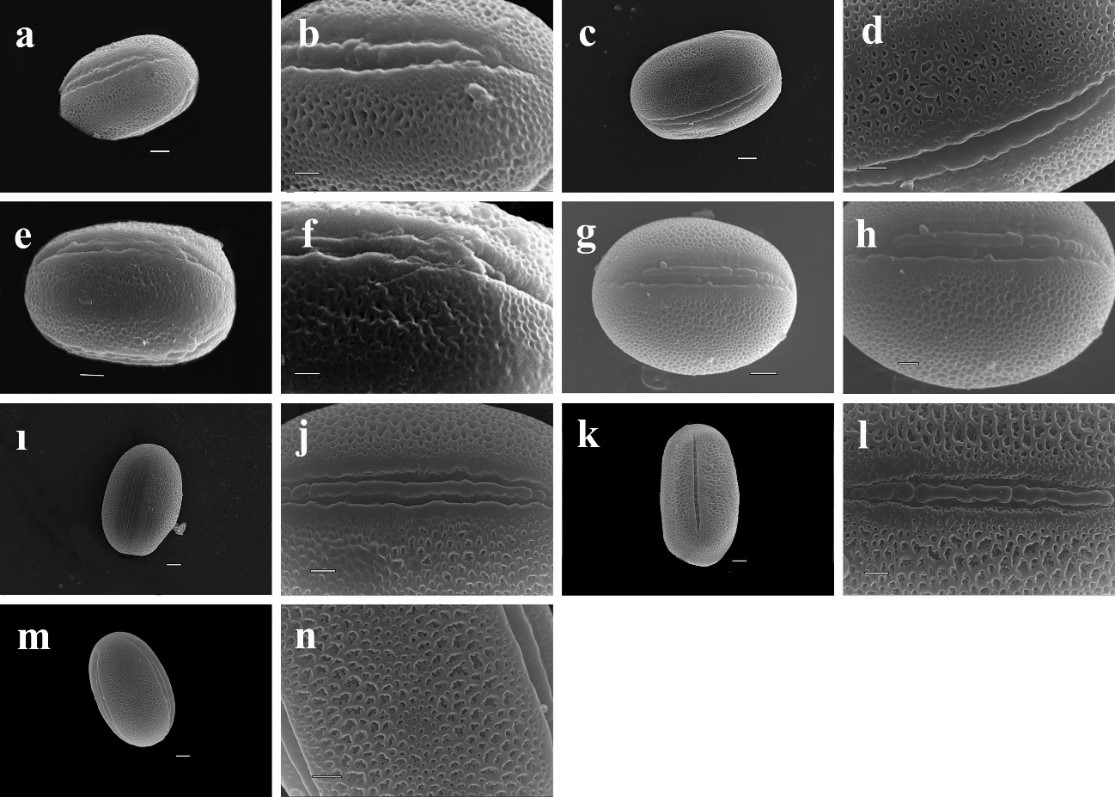
Fig. 3. Electron microphotographs of species of Hedysarum genus of Multicaulia. a-b. H. laxum, c-d. H. leucocladum, e-f. H. huetii, g-h. H. nitidum, ı-j. H. pestalozzae, k-l. H. varium subsp. varium, m-n. H. varium subsp. Syriacum
Numerical analysis of the pollen characters traits
A dendrogram produced with the use of the UPGMA in the cluster analysis, taking into account the ten palynological variables (Tab. 2) found in the section Multicaulia, is shown in Fig. 4. Similarities among the investigated taxa were reflected in the dendrogram, which revealed two main groups: group A (with 45% similarity) contained only H. huetii, which was a distinct species, and group B (with 60% similarity) comprised the remaining 6 taxa. Group B comprised two primary clusters, further denoted as clusters C and D. Two species ( H. pestalozzae and H. varium subsp. varium, with a 73% similarity) made up cluster C. Four taxa with two subgroups, clusters D1 and D2, were included in cluster D. Cluster D1 contained H. laxum and H. leucocladum. Cluster D2 also included 2 species: H. nitidum and H. varium subsp. syriacum.
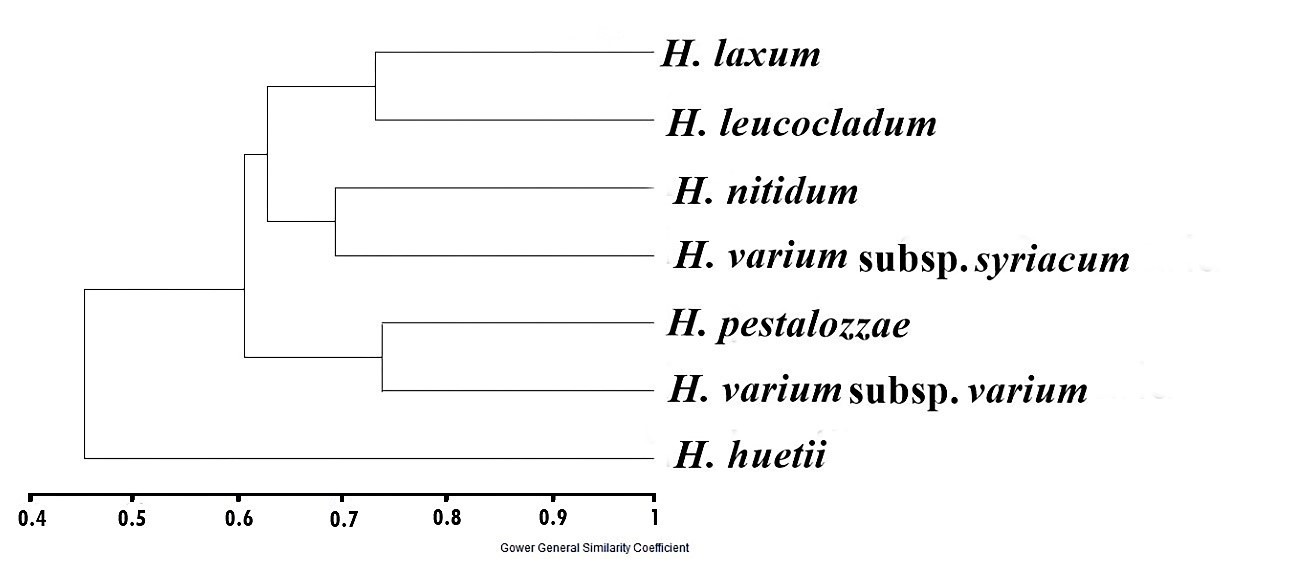
Fig 4. The derived dendrogram based on the palynological characters showing the similarity distance of Hedysarum species.
Discussion
The present study examined the usefulness of using the pollen morphological characteristics of species of the section Multicaulia of the genus Hedysarum in Türkiye. Using these pollen morphological characters, delimitation of taxa was determined.
The findings of current investigation show that the pollen dimensions (E and P/E ratio), the thickness of the exine and intine, the shape of the polar and equatorial views, and the aperture type of the Türkiyeli representatives of Hedysarum were comparatively homogeneous (Tab. 2, Fig. 1). Ohashi (1971), Ferguson and Skvarla (1981), Faegri and Iversen (1989), Moore et al. (1991), Pavlova and Manova (2000), and Ghanavati and Amirabadizadeh (2012) confirmed that the pollen morphological characteristics of the different sections of Hedysarum are essentially the same. Polar and equatorial axes were useful pollen characters to group species with the aid of numerical analysis as mentioned before in Astragalus (Bagheri et al. 2019). Another study declared that Onobrychis genus had oblate-spheroidal pollen shape (Avcı et al. 2013).
All of the species investigated in this research had tricolpate-type pollen grains with microreticulate-perforate exine ornamentation. Only H. varium subsp. varium was different from others, with reticulate-perforate exine. Reticulate and finely reticulate exine ornamentation were reported previously in the genus Hedysarum by Civelek et al. (1999), Pavlova and Manova (2000), and Dural and Citak (2015). The sculpture of exines were determined with three types counted reticulate, suprareticulate and microreticulate in close relatives of Hedysarum genus, Astragalus and Onobrychis (Avcı et al. 2013, Bagheri et al. 2019). Perveen and Qaiser (1998) reported that the ornamentation, and in particular, the shape and size of the muri and lumina, was an important character for Fabaceae (Pavlova and Manova 2000, Ghanavati and Amirabadizadeh 2012, Dural and Citak 2015). The colpus membrane has been identified as being covered with large and small sculptural elements by different researchers in the genus Hedysarum (Pavlova and Manova 2000, Dural and Citak, 2015). Moreover, in this study the same sculptural elements were observed in the subsection Multicaulia as described for the operculum with psilate sculpture.
The UPGMA dendrogram derived based on the palynological characters discriminated the species of the subsection Multicaulia. The positions of the species and the relationships of the subsection Multicaulia of the genus Hedysarum in the cluster were found to be in agreement with the previous classification made on a large scale (Hedge 1979, Hamzaoğlu and Koç 2020). H. huetii, which is a very distinct species, can be easily separated from the others by its fleshy and ovate-orbicular leaflets; hence, it was alone in a different subclade. It was very surprising that H. pestalozzae and H. varium subsp. varium were in the same clade, because of their broad morphological differences, particularly regarding the color of the corolla. In addition, the positions of H. varium subsp. syriacum and H. nitidum in the same clade were surprising due to their morphological differences. This situation raised the question of whether or not new taxonomic regulations for these taxa are required. The last two species, H. laxum and H. leucocladum, which are local and endemic , were in the same subclade. The distribution of the species of the subsection Multicaulia has raised the question of whether or not the relationships of the taxa should be reviewed one more time based on their pollen characteristics. Furthermore, different lumina dimensions, lumina types, and densities among taxa may be associated to their multiple germination requirements or dispersal strategies, which may have developed as a result of an evolutionary adaptation independent of various habitat types or microclimatic regions, particularly for endemics. More data are needed to solve the systematic problems of this section, especially regarding the phylogenetical relationships of the taxa.
Conclusion
To sum up, an in-depth investigation of the pollen characters was highly helpful in classifying the studied taxa. In future work, we propose that the systematic problems still unresolved in Hedysarum taxa should perhaps be solved by combining morphological with anatomical, micromorphological, karyological, and additional molecular data.