
Introduction
The Mediterranean basin is considered a biodiversity hotspot on account of its exceptional concentration of endemic species and the loss of habitats suitable for them (Myers et al. 2000). A significant component of this biodiversity is preserved in the Mediterranean islands due to the presence of steno-endemic taxa and distinctive plant assemblages (Médail 2017, Vargas 2020). The islands of the eastern Adriatic, with its multitude of archipelagos, are particularly important from this point of view, and they are characterised by a very high plant richness, comprising almost 1800 vascular taxa (Nikolić et al. 2008).
The phytosociological investigation of the north-eastern Adriatic using the approach of Braun-Blanquet (Westhoff and van der Maarel 1980) began in the 1930s, with the pioneering studies of Croatian botanist Stjepan Horvatić on some islands, such as Pag and Rab (Trinajstić 2001 and references therein). On those islands, one of the most widespread aspects of vegetation was represented by open Mediterranean-sub-Mediterranean grasslands the particular floristic composition of which justified their attribution to an endemic alliance ( Chrysopogono grylli-Saturejion subspicatae Horvat et Horvatić 1934 nom. inval.). Subsequently, that alliance was classified in an order ( Scorzonero villosae-Chrysopogonetalia grylli Horvatić et Horvat in Horvatić 1963) and class also endemic to the Western Balkans ( Brachypodio-Chrysopogonetea Horvatić 1963) (see Terzi 2011).
According to the current European syntaxonomic framework, the so-called EuroVegChecklist (EVC, Mucina et al. 2016), those grasslands mostly belong to the western Balkan alliances Chrysopogono grylli-Koelerion splendentis and Scorzonerion villosae, and to the trans-Adriatic order Scorzoneretalia villosae, included in turn in the class Festuco-Brometea (but see also Terzi et al. 2023).
The first survey - the subject of Horvatić's doctoral thesis (Trinajstić 2001) - concerned the vegetation of the island of Plavnik, in the Kvarner area (Croatia), where Horvatić (1927) described five grassland syntaxa, namely Festucetum valesiacae, Asphodeletum microcarpi, Helichrysetum italici, “ Helichrysetum salviosum” and Andropogonetum grylli. For each of them, a diagnosis was provided consisting of a variable number (from 5 to 10) of quadrats of 1 square metre each, with a complete list of taxa accompanied by quantitative scores. Although each vegetation type was sampled on only a small area, Horvatić (1927) gave a fairly precise idea of the main dominant and codominant taxa present at that time.
Quite surprisingly, neither Horvatić's later papers (e.g. Horvatić 1963, 1973) nor the subsequent syntaxonomic revisions of Croatian grasslands of the order Scorzoneretalia villosae (e.g. Horvat et al. 1974, Trinajstić 2008, Terzi 2015) mentioned these five syntaxa. Moreover, the herbaceous vegetation of the island of Plavnik has never been studied subsequently.
This island presents a rather interesting situation from a vegetation point of view because it has remained uninhabited for a long period of time, being used as a hunting reserve and only occasionally visited by hunters and tourists (along the coastal areas). The conspicuous presence of animals, sheep and fallow deer, left in the wild, has presumably significantly increased the grazing pressure on the grassland ecosystems.
The present work therefore aims to (i) provide new relevés for the herbaceous vegetation of the island of Plavnik, using a more reliable plot size than that used by Horvatić (1927), (ii) assess the changes in vegetation that have occurred over almost 100 years, (iii) provide a syntaxonomic framework for the grassland associations occurring on the island, and (iv) identify the main ecological drivers of grassland diversity.
Study area
The island of Plavnik (surface area 8.63 km2, coastline length 18.47 km) belongs to the north-eastern Adriatic group of islands in the Kvarner Bay, Croatia (Fig. 1). It is located between the islands of Cres and Krk and it stretches in a northwest-southeast direction for a length of 6.3 km. The coastline of the island is not indented: the northern coast is very steep and poorly accessible, while the southern is flat, gentle, accessible and with several coves.
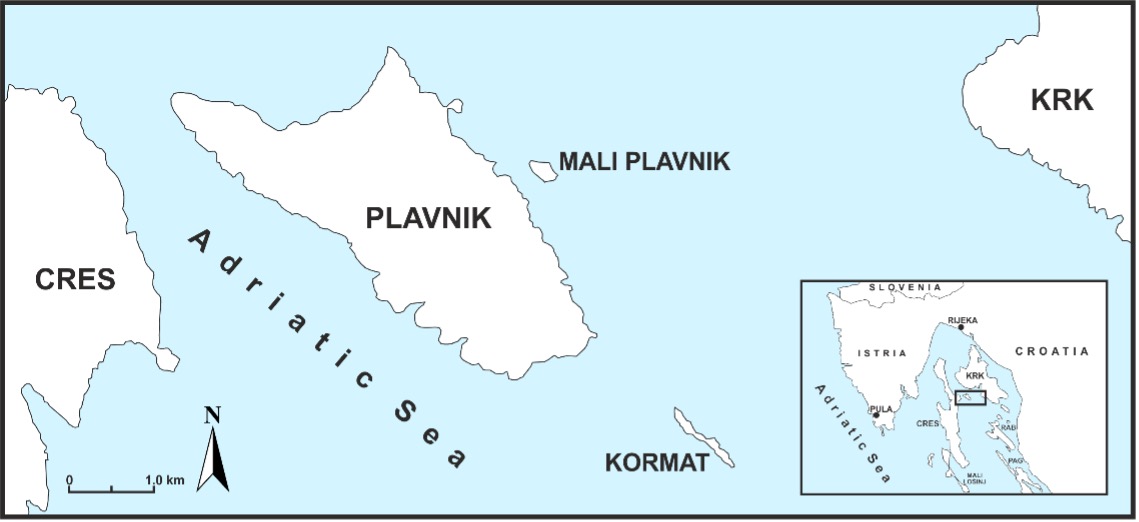
Fig. 1. The island of Plavnik is located between the islands of Cres and Krk in the Kvarner Bay (Croatia). The square on the map in the lower right corner shows the study area in the NE Adriatic context.
The island has high terrain ruggedness. The northern and central part is a karstic plateau with an elevation of approximately 90–180 m a.s.l., bordered by steep rocky slopes exceeding 30°. The highest peak reaches an elevation of 193.6 m a.s.l. To the south-east, the karstic plateau gradually passes into lowlands with altitudes lower than 30 m a.s.l. (Fig. 2).

Fig. 2. Physiognomy of the plant associations on the island of Plavnik in May 2023: A – Helichrysetum italici Horvatić 1927 on the island’s eastern coast, B – Helichrysetum italici salvietosum officinalis subass. nov. covers the open areas on the island’s western slopes, C – Hedypnoido rhagadioloidis-Asphodeletum ramosi ass. nov., in the southernmost part of the island, D – Festuco valesiacae-Poetum bulbosae ass. nov. occurs on the karstic plateau in the north-central part of the island.
The bedrock consists mainly of carbonate rocks (Fuček et al. 2015). The Upper Cretaceous carbonate deposits can be recognized in different types of vertically alternating lithotypes. Multi-layered and massive recrystallized limestones with radiolithid-chondrodotic lithosomes are developed on both western and eastern coasts, while in the central part there are medium-thick layered shallow-sea mudstones and granular limestones with benthic foraminifera, radiolithid rudists and chondrodonts. The brown soils on limestone are predominant on the island (Vukadinović 2021).
The area is influenced by both Mediterranean and continental climates and characterized by intrusions of cold air during the winter and convective precipitation in the summer. These specific conditions result in the lack of a distinct dry period typical of the Mediterranean climate (Šegota and Filipčić 2003). The average annual air temperature is 15.3 °C and precipitation averages 1253.7 mm yr-1 (data from the town of Krk station for 1998-2022, Croatian Meteorological and Hydrological Service). The highest monthly average temperature is 30.5 °C in July, and the lowest 2.5 °C in January. The absolute maximum of 38.8 °C was recorded on 21 July 2015, and the absolute minimum (-9.0 °C) on 24 January 2006. The greatest rainfall comes in September, October and November with averages of 155.6, 162.0 and 178.0 mm, respectively. In the period from June to August the total rainfall is 184.2 mm. Northern winds (NW, ENE, NE) prevail throughout the year. This area has 2521 hours of sunshine per year (data for the island of Rab station for 1998-2022). On average the relative air humidity is 70%.
Phytoclimatic indices were calculated according to Rivas-Martínez et al. (2011): annual positive temperature (Tp) is 1,834; continentality index (Ic) is 18.9; thermicity index (It) is 238; ombrothermic index (Io) is 0.68. According to these indices, the neighbouring Krk station lies within the temperate macrobioclimate, meso-temperate thermotype and semiarid ombrotype.
The observed global and regional warming has been unequivocal since the middle of the 20th century (IPCC 2014) and is present along the eastern Adriatic coast (Branković et al. 2013). Climate change shows a tendency to manifest in drier conditions for the Adriatic (Patarčić et al. 2014, WBG 2021), which is a common feature for the broader European part of the Mediterranean (EEA 2019). However, for example, the precipitation change is regionally highly variable and further analyses are needed to define the assessment of climate change impacts at the regional and local scale (Gajić-Čapka et al. 2018).
From the phytogeographic point of view, the eastern and southwestern part of the island is covered by stands of the Fraxino orni -Ostryion alliance i.e. Querco pubescenti- Carpinetum orientalis Horvatić 1939 association. The meso-Mediterranean evergreen vegetation of the Fraxino orni-Quercion ilicis alliance is developed only in the northwestern part of the island. However, woodlands have been significantly changed due to the long-term anthropogenic influences and nowdays only degraded vegetation types, i.e. maquis, garrigue and grasslands, may be found. Large areas of trees were felled between the two world wars to obtain open pastures, intended for sheep rearing. The island is classified among Croatia's Important Plant Areas (IPAs) and is part of the Natura 2000 ecological network (Topić and Vuković 2010, Official Gazette 2019).
The island has had no permanent residents or settlements during the last centuries. Between the two world wars, only a few herding groups lived on the island to provide living conditions for livestock, mainly sheep, while goats made up only 10-20% of the total number of head (Dumančić 1988). After the Second World War, such a simple nomadic way of life gradually faded away. The sheep population has drastically decreased, while goats, in general, have practically disappeared on the eastern coast of the Adriatic. There are no available data on the number of sheep in Plavnik over last century. In addition, the non-native European fallow deer ( Dama dama) was introduced in 1995. According to the data of the local hunting association Farma Plavnik d.o.o. (personal communication), a number of the fallow deer, with regular annual shooting of ca. 50 nap, are stable and amounts to about 160 nap. The maximum nutritional requirements for females are during lactation, and this period coincides with the maximum development of vegetation.
Vegetation sampling and data analysis
The map of the distribution areas of the five types of plant communities on the island of Plavnik provided by Horvatić (1927: p. 44) was compared with the most recent satellite images available on Google Earth Engine, to identify areas still covered by herbaceous vegetation. Those areas, and in particular the locations mentioned in that paper, were surveyed. The vegetation types were identified in the field based on their dominant and codominant taxa. Only for the “ Andropogonetum grylli”, whose dominant taxon should have been Chrysopogon gryllus, we were unable to find any phytocoenosis clearly dominated by this plant, which is nowadays rather rare on the island.
The remaining four vegetation types were sampled through 29 relevés, made in May 2023 according to the phytosociological approach (see Westhoff and van der Maarel 1980). The relevés were sampled in quadrats of 25 m2 (5 m × 5 m). All vascular plants in the plots were recorded and their cover-abundance values were visually estimated using the Braun-Blanquet cover-abundance scale with the subdivisions 2m, 2a and 2b as proposed by Barkman, Doing and Segal (in: van der Maarel 1979). This extended Braun-Blanquet scale (i.e. r, +, 1, 2m, 2a, 2b, 3, 4, 5) was replaced by the 9-point ordinal scale (i.e. 1-9, see Westhoff and van der Maarel 1980), before the statistical analyses. The main physical features of the plots (e.g., geographic coordinates, slope, exposure, vegetation cover, rockiness, and stoniness) were also recorded. Stoniness is defined here as the percentage of plot coverage by stones (incoherent material), regardless of their size. Rockiness, on the other hand, indicates the percentage of coverage of the plot determined by the outcropping rocky substrate.
The data matrix (29 relevés × 115 taxa) was subjected to hierarchical clustering using Ward's method on a Chord distance matrix (i.e., Euclidean distance on pre-transformed data through normalization by sites). Four interpretable clusters were subjectively defined by visual examination of the dendrogram.
To visualize the floristic and ecological relationships among the four clusters of relevés, nonmetric multidimensional scale (NMS) ordination was performed by using the Chord distance. Both cluster analysis and ordination were performed through the PcOrd 6.22 software package (McCune and Mefford 2011), the NMS with the "slow and thorough" option of the autopilot mode.
To assess ecological differences among the four plant communities, taxa score-weighted means of the Ellenberg-type Indicator Values (IVs) of temperature (T), soil reaction (R), moisture, and nutrient content were calculated for each relevé, based on the EIVs provided for Italy - which is the closest country geographically and ecologically to the study area - in Tichý et al. (2023).
In addition, given the presence of high numbers of sheep and fallow deer on the island of Plavnik and the possible importance of the disturbance regime in shaping the different plant communities, three additional disturbance-related IVs, taken from Midolo et al. (2023), were considered. In more detail, the following IVs, were calculated on taxa score-weighted means for each relevé: Disturbance severity (Ds), Disturbance frequency (Df), and Grazing pressure (Gr). Joint plots were used to represent linear correlations between IVs and ordination scores, with an r2 cut-off of 0.3.
Indicator species analysis (ISA) (Dufrêne and Legendre 1997) was used to identify the indicator taxa of the four main clusters of relevés obtained by cluster analysis, and their combinations in larger groups (see De Caceres et al. 2010). The indicator values were calculated for all taxa present in at least three relevés. The statistical significance (P < 0.01) of the indicator values was tested by a Monte Carlo test with 10000 permutations. The ISA was run in R software (R Core Team 2023) through the package indicspecies 1.7.12 (De Cáceres and Legendre 2009).
In order to verify whether the relevés from the island of Plavnik in which Asphodelus ramosus was dominant (i.e. cluster 'He' in Figs. 3 and 4) could represent an association independent of the others described so far in the eastern Adriatic, we compared these relevés with the associations Bromo erecti-Chrysopogonetum grylli Horvatić 1934 and Narcisso tazettae-Asphodeletum microcarpi Šegulja 1969. In more detail, a second data matrix (55 relevés × 283 taxa) was obtained by adding the seven relevés reported in Table 3 to the other 48 relevés carried out by i) Horvatić (1934, rels. 11-23 of table XXVIII) on the island of Pag, Croatia; ii) Stanišić-Vujačić et al. (2022, rels. 1-17 of table 1) at Ćemovsko polje, Montenegro, iii) Šegulja (1970, rels. 1-8 of table II) in Labin, Istria, Croatia, and iv) Hećimović (1984, Narcisso tazettae-Asphodeletum microcarpi sisymbrietosum officinalis Hecimović 1984, rels. 1-10 of table 5) on the islets of Bobara and Mrkan, in southern Croatia.
Using the same methods as described above, an NMS ordination was performed on that matrix to visualise the floristic relationships between the relevés, and then an indicator species analysis was run to identify the taxa that distinguish the Asphodelus ramosus community on Plavnik Island from those elsewhere.
The taxonomic nomenclature follows Euro+Med (2006-2023). Bromopsis erecta and B. condensata were treated under the same tag, the latter being more abundant and frequent on the island of Plavnik.
Syntaxonomic nomenclature of alliances, orders and classes follows the EuroVegChecklist (https://floraveg.eu/vegetation/, retrieved on 10 October 2023, as modified by Terzi et al. 2024a), whose correct names are given without author citations. Syntaxon names not included in the EVC, but are instead provided with author citations. Names reported exactly as mentioned in the original papers are reported in quotation marks.
Results
The dendrogram (Fig. 3) and further NMS (Fig. 4) showed four main clusters of relevés corresponding to the plant communities dominated by (Fe) Festuca valesiaca and Poa bulbosa, (As) Asphodelus ramosus, (He) Helichrysum italicum, and (Sa) Salvia officinalis and Helichrysum italicum.
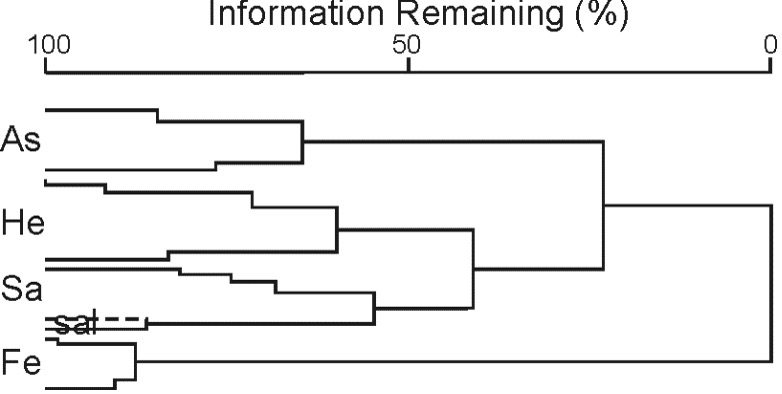
Fig. 3. Hierarchical clustering (Ward's method, Chord distance) shows four main groups of relevés corresponding to the plant communities: As – Hedypnoido rhagadioloidis-Asphodeletum ramosi, He – Helichrysetum italici, Sa – Helichrysetum italici salvietosum officinalis, “sal” indicates a group of relevés clearly dominated by Salvia officinalis, Fe – Festuco valesiacae-Poetum bulbosae.
The cluster Fe groups together relevés from the summit plateau of the island, at more than 150 m a.s.l., where Horvatić (1927) had previously described the association Festucetum valesiacae. The mean number of taxa is 33 whereas in all the relevés of this cluster 53 taxa were found. In addition to F. valesiaca and P. bulbosa many species were recorded with high frequency (see Tab. 1). The indicator species associated to this cluster are Trifolium suffocatum, Erodium cicutarium, Crepis neglecta subsp. neglecta, Petrorhagia saxifraga, and Marrubium incanum.
Tab. 1. Festuco valesiacae-Poetum bulbosae ass. nov. on the island of Plavnik. Life-forms (LF): Ch – chamaephytes, G – geophytes, H – hemicryptophytes, P – phanerophytes, T – therophytes.
The second partitioning level separates the cluster As. This cluster represents the vegetation clearly dominated by Asphodelus ramosus, the latter being always recorded with a cover greater than 75%, except in one relevé. This vegetation type is well represented in the southern part of the island. The indicator taxa of this cluster are Asphodelus ramosus, Hedypnois rhagadioloides, Crepis zacintha, and Centaurium maritimum. The mean number of taxa per relevé is 32.
The third cluster He represents the most widespread type on the island: open vegetation dominated by Helichrysum italicum and Koeleria splendens, and with other taxa typical of the Festuco-Brometea class occurring with high frequency (e.g. Bromopsis erecta, Euphorbia cyparissias, Tab. 2).
Tab. 2. Helichrysetum italici typicum (rels. 1-10) and Helichrysetum italici salvietosum officinalis (rels. 11-14); transition to Stipo-Salvietum officinalis (rels. 15-16) on the island of Plavnik. For abbreviations of Life-forms [LF], see caption of Tab. 1.
The indicator species of this cluster are Reichardia picroides, Bupleurum veronense, Plantago holosteum, and Hippocrepis comosa. This vegetation type usually develops on rocky and stony ground whereas on steep slopes on the eastern part of the island, in some places, it gives way to a Salvia officinalis- Helichrysum italicum community, represented by the cluster Sa. The latter corresponds to the “ Helichrysetum salviosum” described by Horvatić (1927) and includes seven relevés, with a mean number of taxa of 21. This cluster is actually heterogeneous with some relevés which represent a variant of the Helichrysetum italici due to the presence and co-dominance of both Helichrysum italicum and Salvia officinalis, whereas two relevés (indicated as “sal” in Figs. 3 and 4) are clearly dominated by the latter. Salvia officinalis is the only indicator species of this cluster.
The NMS (Fig. 4) resulted in a two-axis solution, with a final stress 8.96, indicating a fairly good ordination.
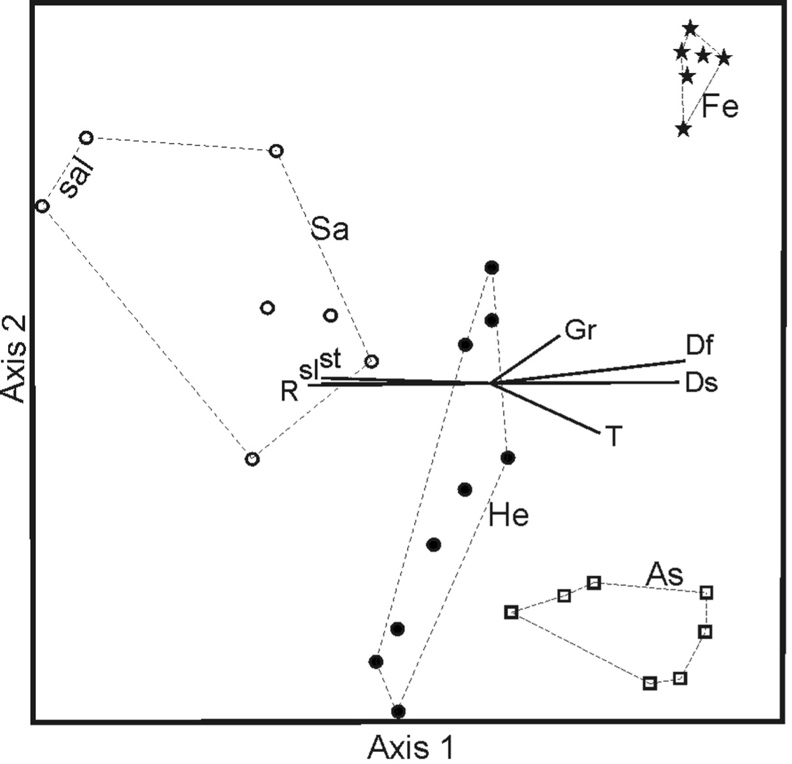
Fig. 4. Non-metric multidimensional scaling of releves in the dataset. Abbreviations: As – Hedypnoido rhagadioloidis-Asphodeletum ramosi, He – Helichrysetum italici, Sa – Helichrysetum italici salvietosum officinalis, “sal” indicates a group of relevés clearly dominated by Salvia officinalis, Fe – Festuco valesiacae-Poetum bulbosae. Ds – disturbance severity, Df – disturbance frequency, Gr – grazing pressure, T – temperature, R – soil reaction, st – stoniness, sl – slope.
The two axes accounted for 79.5 of the total variation (axis 1 = 65.3%, axis 2 = 14.2%). The four clusters occupy different parts of the ordination space without overlapping each other. Axis 1 separates the clusters He and Sa among them and from Fe and As. This axis is positively correlated with Df, Ds, Gr and temperature whereas it is negatively correlated with slope, stoniness and reaction. The correlation strengths of the EIVs of moisture and nutrient content with the ordination axes were lower than the selected r2 cut-off and we will not comment on those variables further. Therefore, the Sa cluster groups relevés taken on steeper slopes than the other clusters and with high percentages of stones. The Fe and As clusters have the highest values of disturbance, in terms of both severity and frequency. The lowest reaction values for these two clusters are probably related to disturbance. Axis 2 clearly separates Fe from As, the former with greater values of grazing, the second with greater values of temperature, being in the southern part of the island at the lowest altitude and mostly on the south-facing side.
The NMS ordination of the relevés of the As cluster together with those of the associations Bromo erecti-Chrysopogonetum grylli and Narcisso tazettae-Asphodeletum microcarpi resulted in a three-axis solution with a final stress of 7.5 (Fig. 5).
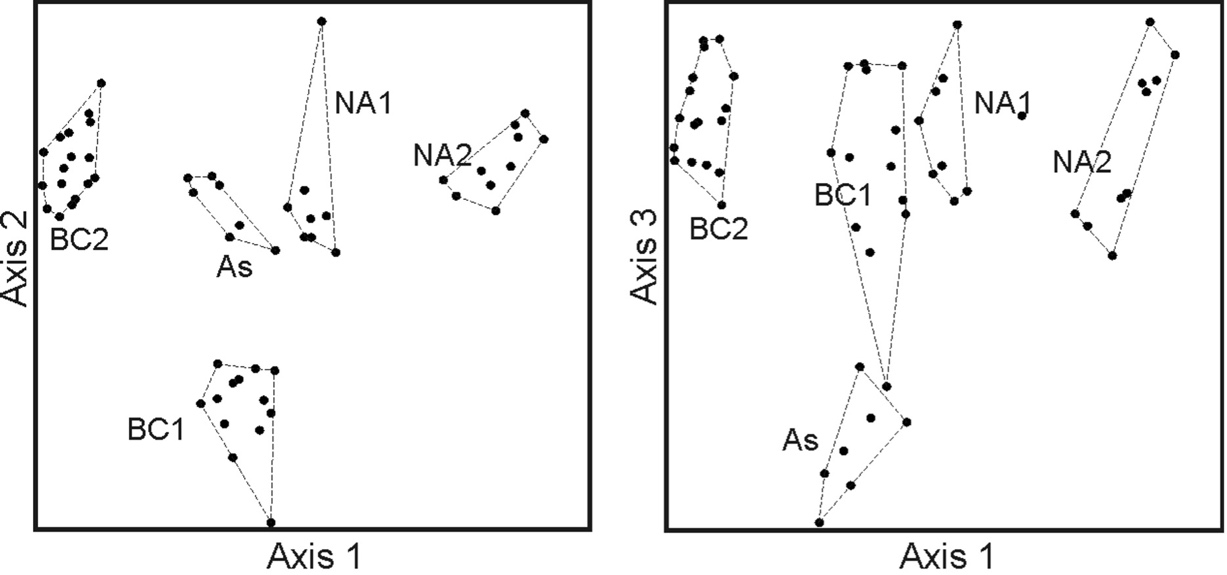
Fig. 5. Non-metric multidimensional scaling (with axes 1, 2 and 3) of eastern Adriatic associations with Asphodelus ramosus. Abbreviations: HA – Hedypnoido rhagadioloidis-Asphodeletum ramosi, BC – Bromo erecti-Chrysopogonetum grylli (BC1 from island of Pag, Croatia, and BC2 from Montenegro), NA – Narcisso tazettae-Asphodeletum microcarpi (NA1 from Istria and NA2 from islands of Bobara and Mrkan, Croatia).
Most of the variance is represented by axis 1 (55.1 %) whereas axes 2 and 3 account for 22.6 % and 2.5 %, respectively. The relevés of the two associtions Bromo erecti-Chrysopogonetum grylli (BC1 and BC2 in Fig. 5) and Narcisso tazettae-Asphodeletum microcarpi (NA1 and NA2) are separated on axis 1, whereas the relevés of the cluster As are in the middle. The latter are clearly separated from the others on axis 3 (Fig. 5). Interestingly, the relevés of the two phytosociological tables of the Bromo erecti-Chrysopogonetum grylli (i.e. BC1 and BC2) and those of the two tables of the Narcisso tazettae-Asphodeletum microcarpi (i.e. NA1 and NA2) form four well separated groups of relevés.
The cluster As was found to be well distinguished in the species-space from the other associations with which it has no overlap. The indicator species analysis indicated the following 11 taxa as diagnostic for the As cluster (indicator values in brackets): Hedypnois rhagadioloides (74), Linum trigynum (72.3), Galium murale (70.6), Euphorbia cyparissias (68.9), Geranium purpureum (67.2), Sideritis romana (62.4), Vulpia ciliata (60.8), Centaurium maritimum (56.3), Plantago bellardii (56.3), Scorpiurus muricatus (56.3), and Lotus hispidus (42.3).
Discussion
A direct quantitative comparison of Horvatić's data with the new data is not completely possible because the plot sizes used in the two surveys are too different (see also Dengler et al. 2009). For example, the five plots used by Horvatić (1927) to represent the Festucetum valesiacae sample a total area of 5 m2 and include 17 taxa. The new dataset comprises six relevés of the same area - each with a plot size of 25 m2 and containing an average of 33 taxa - for a total number of taxa of 53. Furthermore, the quantitative values of the taxa in Horvatić's article are estimated on a scale comprising at least 10 (undefined) values (-, -1, 1, 1-2, 2, 2-3, 3, 3-4, 4, 5), which are not directly comparable with the 9-points scale used in this paper. However, on the basis of qualitative considerations, it can be deduced whether the plant communities remained fairly similar or not during the 100 years.
As written above, although Horvatić (1927) described five grassland syntaxa on the island of Plavnik, we were only able to find four grassland community types. In fact, the area originally covered by Andropogonetum grylli is completely occupied by a plant community dominated by Asphodelus ramosus, whereas Chrysopogon gryllus - i.e. the dominant taxon of the Andropogonetum grylli - is quite rare on the island.
Festuco valesiacae-Poetum bulbosae
The Festucetum valesiacae described by Horvatić (1927) for the continuous herbaceous expanses on the summit of the island is no longer present. This association was dominated by Festuca valesiaca, followed as cover by Bromopsis erecta and Plantago holosteum, and included several other species of the class Festuco-Brometea class (e.g. Thymus longicaulis, Chrysopogon gryllus, Koeleria splendens), to which it clearly belonged. In that area, the physiognomic situation appears quite different nowadays, with herbaceous phytocoenoses interspersed with shrubs and trees of Juniperus oxycedrus, generally on flat surfaces, where sheep and fallow deer find shelter (Fig. 2d). The herbaceous vegetation is dominated by Poa bulbosa and F. valesiaca, followed by some Trifolium species ( T. campestre, T. suffocatum, T. subterraneum, T. scabrum), and Koeleria splendens. The results show that this plant community is clearly distinct from the others in terms of floristic composition and ecology. Indeed, it is the most disturbed in terms of intensity and frequency of disturbance (Ds and Df in Fig. 3) together with Asphodeletum ramosi but, unlike the latter, it has a higher grazing pressure (Gr). Therefore, the plant community dominated by Poa bulbosa and Festuca valesiaca (Fe in Fig. 3) is here considered to be a new association, the Festuco valesiacae-Poetum bulbosae ass. nov. hoc loco (holotypus rel. 1 in Tab. 1), possibly derived from Festucetum valesiacae - of which Festuca valesiaca, Koeleria splendens, and other taxa are still present - as a consequence of the prolonged stay of sheep and deer and intensive grazing.
The presence with high cover and/or frequency of Poa bulbosa, Trifolium suffocatum, T. subterraneum, Erodium cicutarium, Plantago lanceolata, and other species, indicates that the Festuco valesiacae-Poetum bulbosae belongs to the Poetea bulbosae, i.e. a class representing heavily grazed pastures, which are mainly present in the western Mediterranean basin (e.g. Rivas Goday and Rivas-Martínez 1963). This class comprises a single order, Poetalia bulbosae, and several alliances, none of which occur in Croatia (Preislerová et al. 2022), with the exception of the Romuleion (Škvorc et al. 2017). This alliance was originally invalidly described by Oberdorfer (1954, see Terzi et al. 2024b) for the south and southeast of the Balkan Peninsula, and was identified by 20 character species, none of which are present in our relevés (at least in the phenological stage of vegetation at the time of sampling). The alliance was originally classified in the class “ Thero-Brachypodietea” (Oberdorfer 1954), and then moved to the “ Brachypodio-Chrysopogonetea” (Horvat et al. 1974), “ Saginetea maritimae” (Rodwell et al. 2002), and Tuberarietea guttatae (Čarni et al. 2014). The EVC has instead classified the Romuleion in the Poetea bulbosae (EVC) to represent “ Macedonian seasonal perennial pastures on acidic substrates”. The Poetea bulbosae, indeed, are most frequently found on acid substrates (Oberdorfer 1954, Čarni et al. 2014).
The numerous taxa of the Stipo-Trachynietea distachyae found in the Festuco valesiacae-Poetum bulbosae are linked to the basic substratum of the island of Plavnik, while those of the Tuberarietea guttatae highlight the dynamic relationships with the oligotrophic grasslands of this class, where the Romuleion was framed by some authors (e.g. Čarni et al. 2014). Two associations of the Poetalia bulbosae were recently described near Podgorica, Montenegro, on fluvioglacial deposits and calcareous rocky substrate and framed in the Romuleion (Stanišić-Vujačić et al. 2023). The floristic composition of those associations, however, includes only three character species of this alliance ( Romulea linaresii subsp. graeca, and the very rare Hypochaeris cretensis and Hedypnois rhagadioloides). The associations from Montenegro were found in an area where rocky grasslands of the alliance Chrysopogono grylli-Koelerion splendentis are widespread and indeed at least one of those associations ( Romuleo bulbocodii-Poetum bulbosae Stanišić-Vujačić et al. 2023) is characterized by the presence of numerous Festuco-Brometea species (Stanišić-Vujačić et al. 2022, 2023). The Festuco valesiacae-Poetum bulbosae, derived from the Festucetum valesiacae, also has dynamic relationships with that alliance and in fact shares several species (e.g., Koeleria splendens, Thymus longicaulis) with the Montenegrin associations. However, the natural potential vegetation of the Plavnik area is different from that of the surrounding of Podgorica, in Montenegro.
The floristic relationships of the Festuco valesiacae-Poetum bulbosae with the Romuleion are, however, even weaker, given that none of the character species of this alliance were detected in our relevés. For this reason, the Festuco valesiacae-Poetum bulbosae is only provisionally classified in the Romuleion, pending the expansion of knowledge about the variability of the Poetalia bulbosae in the Balkans by new phytosociological studies. It is indeed plausible to admit that a second alliance of the Poetalia bulbosae, in addition to the Romuleion, is present in the Western Balkans and that it is dynamically linked to a degradation of the grasslands of the Scorzoneretalia villosae (not present in Greece, where the Romuleion was originally described), but a large-scale comparison is needed to confirm this hypothesis, which is beyond the scope of this paper. In any case, the Festuco valesiacae-Poetum bulbosae documents for the first time, based on phytosociological data, the presence of Poetalia bulbosae and Poetea bulbosae in Croatia.
Helichrysetum italici
The most frequent vegetation type on the island is represented by the Helichrysum italicum dominated plant community (“He” in Figs. 3 and 4), which grows on rocky and stony substrates throughout the island. Helichrysum italicum is widespread in southern Europe with several subspecies and the nominal one, Helichrysum italicum subsp. italicum, which is the only one found on the island of Plavnik, has a range in Europe that includes southern France, Italy and the Balkans (Euro+Med 2023). This subspecies grows in different types of vegetation, such as thermophilic scree communities of the Scrophulario-Helichrysetalia (Brullo and Spampinato 1991), and coastal sub-aerohaline dwarf scrub vegetation of the Helichrysetalia italici (Foucault 2020). In the Balkans, it has been considered a character taxon of the “ Brachypodio-Chrysopogonetea” (Horvatić 1963, Horvat et al 1974). Helichrysum italicum is a suffruticose chamaephyte and, where it is dominant, the vegetation physiognomy takes on the appearance of a chamaephytic garrigue (Horvatić 1927). In the Dinaric Arc mountain range, this type of vegetation is often rich in many species typical of grasslands, so that it is not always possible to distinguish chamaephytic garrigues from 'pure' grasslands from a syntaxonomic point of view. In fact, the alliance Chrysopogono grylli-Koelerion splendentis, of the order Scorzoneretalia villosae, includes vegetation types dominated by both hemicryptophytes and dwarf chamaephytes, such as Helichrysum italicum and Salvia officinalis.
The same situation can be observed on the island of Plavnik where the Helichrysetum italici includes Helichrysum italicum, as the dominant, together with many grassland taxa, such as Bromopsis condensata, Trifolium campestre, Dactylis glomerata subsp. hispanica, Salvia pratensis, Euphorbia cyparissias, Koeleria splendens, Hippocrepis comosa. This plant community grows on incoherent substrata with a floristic composition that resembles that sampled by Horvatić (1927) a hundred years ago, except for the absence of Chrysopogon gryllus, which has evidently become rather rare on the island (Fig. 2a). Although the total number of species found in the new relevés is far higher than that recorded by Horvatić (1927), this difference is almost certainly due to the different sizes of the plots used in the two sampling surveys. The species that shape the physiognomy of the Helichrysetum italici are Helichrysum italicum, Bromopsis condensata, Koeleria splendens, and Trifolium campestre. The association clearly belongs to the Chrysopogono grylli-Koelerion splendentis due to the presence of characteristic species of the class Festuco-Brometea (e.g. Bromopsis condensata, Euphorbia cyparissias, Linum tenuifolium, Festuca valesiaca, Hippocrepis comosa), of the order Scorzoneretalia villosae and of the alliance (e.g. Thymus longicaulis, Centaurea tommasinii, Plantago holosteum, Bupleurum veronense) (see Terzi 2015). Although Helichrysum italicum is quite common in other associations of this alliance, the specific floristic composition of the Helichrysetum italici is different from that of all the other associations already described. In any case, even if this association was considered as a syntaxonomic synonym of another association belonging to the alliance, the name Helichrysetum italici would retain priority over names described later.
The Helichrysetum italici occurs in its typical aspect (subass. typicum, represented by rels. 1-9, in Tab. 2) on most of the island, and only on some spots does it associate with Salvia officinalis to give rise to the subassociation described in the next section. The typical aspect corresponds to that described by Horvatić (1927) with 10 plots of 1 square meter each. The size of these individual plots thus appears to be much smaller than that normally used to sample the plant communities of the Scorzoneretalia villosae, which varies in 90% of cases between 10 and 150 square meters (Terzi 2015). Because of the small area of the plots sampled by Horvatić (1927), and perhaps also because of the omission of some species, each of those plots counts only a very limited number of taxa. Moreover, the quantitative values associated with these taxa are on a scale hardly comparable with those routinely used in modern phytosociological studies.
Although none of the 10 relevés originally published by Horvatić (1927), if taken alone, can be considered a representative relevé of the Helichrysetum italici for the reasons expressed above, the association is nevertheless validly described because at least the synoptic column of the association, i.e. the last column of table IV (“Skrizaljka IV”, on p. 33, in Horvatić 1927), constitutes a sufficient original diagnosis according to Article 7 of the International Code of Phytosociological Nomenclature (ICPN, Theurillat et al. 2021).
Therefore, we do not designate a lectotype, which would be incomplete and unrepresentative of the typical aspect of the association. Thus, not having a suitable relevé of the original diagnosis to be selected as lectotype of the association (see also Def. VIII of the ICPN), we find it more correct and useful to designate the relevé 3 in Tab. 2 as neotypus hoc loco of the Helichrysetum italici and of its subassociation typicum.
Helichrysetum italici salvietosum officinalis
Along some steep, mainly north-facing slopes on incoherent substrates or on limestone outcrops, the typical aspect of the Helichrysetum italici gives way to another type of vegetation where Salvia officinalis becomes dominant or co-dominant with Helichrysum italicum (Fig. 2b). This aspect was erroneously described by Horvatić (1927) as “Helichrysetum salviosum” (Art. 3e of the ICPN, Theurillat et al. 2021), to indicate a subtype of the Helichrysetum italici. Its floristic composition indeed retains many species of the Helichrysetum italici with the addition of a few others, including the dominant Salvia officinalis, thus forming the subassociation Helichrysetum italici salvietosum officinalis subass. nov. hoc loco (holotypus rel. 13 of Tab. 2). This subassociation has fewer species than the typical one.
The grassland vegetation of the Western Balkans dominated by Salvia officinalis belongs to different vegetation types, mainly framed in the alliances Chrysopogono grylli-Koelerion splendentis and Saturejion subspicatae (Terzi et al. 2023). In our case, the Helichrysetum italici salvietosum officinalis definitely belongs to the first alliance and is perhaps transitional to the Stipo-Salvietum officinalis, whose nomenclatural type was recorded on the island of Unije, also located in Kvarner, about 50 km (as the crow flies) from Plavnik. In particular, two relevés could belong to this association (subcluster 'sal' in Fig. 3), which show the lowest number of taxa, the clear dominance of Salvia officinalis, and the presence of Helichrysum italicum with very low coverage values.
Hedypnoido rhagadioloidis-Asphodeletum ramosi
The vegetation with Asphodelus ramosus develops mainly in the southern part of the island, with large and surprising expanses absolutely dominated by this species, with a cover usually greater than 90% (Fig. 2c). Two relevés recorded in the vicinity of the Helichrysetum italici distribution area represent a transition towards this association and indeed there Helichrysum italicum is the second dominant species with a cover of 25-50%. A very different situation is described by Horvatić (1927) who reports local patches of Asphodeletum microcarpi interspersed in a matrix of other associations ( Andropogonetum grylli and Helichrysetum italici) and sharing several species with them. He notes that this sharing of species between the Asphodeletum microcarpi and the other associations is even more evident during the winter, when Asphodelus ramosus, the main dominant species, survives belowground. In the plots sampled by Horvatić (1927), Asphodelus ramosus was recorded with a score of “2” (or “3” in only one case) together with other species with similar or greater cover, so that some of its plots look like local variants of the Andropogonetum grylli.
On the other hand, as written above, during our surveys on the island we found few plants of Chrysopogon gryllus for which it was not possible to identify the Andropogonetum grylli nor transitions between this association and the Asphodeletum microcarpi. In the most typical situation, Asphodelus ramosus is by far the dominant species, and with the exception of the cases representing a transition with the Helichrysetum italici (rels. 5 and 6 in Tab. 3), in all other relevés the second dominant species never scored higher than 2a (i.e. up to 12% cover).
Tab. 3. Hedypnoido rhagadioloidis-Asphodeletum ramosi ass. nov. on the island of Plavnik. For abbreviations of Life-forms [LF], see caption of Tab. 1.
This plant community comprises numerous annual taxa of the classes Stipo-Trachynietea distachyae and Tuberarietea guttatae. The presence of so many annual plants is not a novelty for this kind of vegetation and has been observed in many other areas of the Mediterranean (see Terzi 2023 and references therein). The plant community dominated by Asphodelus ramosus was found to be the most disturbed grassland type on the island, together with the Festuco valesiacae-Poetum bulbosae. The thriving of Asphodelus ramosus is usually associated with intense grazing pressure and frequent fires. These factors might also be the ecological drivers on the island of Plavnik, where we hypothesise that at least one of them (grazing or fires) has presumably occurred on the island with greater intensity and/or frequency than was the case 100 years ago. Increased grazing pressure is plausible due to the high number of sheep easily observed on that side of the island, while it is more difficult to assess the role of fires as data on their intensity and frequency are not available.
Thus, the present plant community dominated by Asphodelus ramosus on Plavnik Island differs both floristically and ecologically from the Asphodeletum microcarpi described by Horvatić (1927). It is also clearly different from the Bromo erecti-Chrysopogonetum grylli (i.e. Asphodelo-Chrysopogonetum grylli Horvatić 1963, cf. Terzi 2015) and Narcisso tazettae-Asphodeletum microcarpi. In this case, the Plavnik plant community differs in physiognomy, given the clear dominance of Asphodelus ramosus, and floristic composition (Fig. 5). The indicator species that distinguish the plant community of Plavnik Island from those of other locations in the eastern Adriatic are mainly annual species, some of which indicate a slightly acidic to almost neutral substrate (e.g. Centaurium maritimum, Linum trigynum), as is to be expected in heavily grazed karst areas.
In view of the above, the plant community dominated by Asphodelus ramosus on the island of Plavnik is considered here a new association: Hedypnoido rhagadioloidis-Asphodeletum ramosi ass. nov. hoc loco (holotypus rel. 2 of Tab. 3). However, its classification at a higher syntaxonomic rank is rather difficult (see Terzi 2023 and references therein). Biondi et al. (2016) proposed a new class, order and alliance to classify the vegetation dominated by Asphodelus ramosus (and other geophytes) but they have not been accepted because a comprehensive syntaxonomic review on the subject is still lacking (Biurrun and Willner 2020, Terzi 2023). In Montenegro, Asphodelus ramosus is included within communities of the Artemisietea vulgaris, although it is most abundant in the Bromo erecti-Chrysopogonetum grylli (Stanišić-Vujačić et al. 2022). As a result of this as yet undefined syntaxonomic framework, we refrain from classifying the new association into one of the accepted classes.
With the exception of Festucetum valesiacae, Andropogonetum grylli and Asphodeletum microcarpi, which can be considered extinct, the other plant communities on Plavnik Island can be classified as in the following syntaxonomic scheme:
Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947
Scorzoneretalia villosae Kovačević 1959
Chrysopogono grylli-Koelerion splendentis Horvatić 1973
Helichrysetum italici Horvatić 1927 typicum
Helichrysetum italici salvietosum officinalis subass. nov. hoc loco
Poetea bulbosae Rivas Goday et Rivas-Mart. in Rivas-Mart. 1978
Poetalia bulbosae Rivas Goday et Rivas-Mart. in Rivas Goday et Ladero 1970
Romuleion Oberd. 1954 nom. inavl. (?)
Festuco valesiacae-Poetum bulbosae ass. nov. hoc loco
Class: ?
Hedypnoido rhagadioloidis-Asphodeletum ramosi ass. nov. hoc loco
Acknowledgements
The authors thank Steve Latham (United Kingdom) for improving the English. We thank the Croatian Geological Survey for providing the Geological Map of the Republic of Croatia (sheet Cres and Lošinj). We aslo thank the Croatian Meteorological and Hydrological Service for providing climatological data from the meteorological station in the town of Krk, island of Krk, for the period 1998-2022. We would also like to thank two anonymous reviewers for their helpful suggestions.