
Introduction
Coastal dunes are characterized by high ecological diversity, environmental heterogeneity, and variability of species (Martinez and Psuty 2005), as well as by a close interaction between biotic and abiotic factors (Rucco et al. 2014). Vegetation in sand dunes is also influenced by strong seasonality, with two distinct periods, cold rainy winters and dry hot summers, the latter characterized by high irradiance and little or no precipitation. Consequently, plants experience intense stress during summer due to drought, large evaporative demand and high irradiance at high temperatures (Antunes et al. 2018). The ecological conditions of coasts are often stressful or even critical, with their flora growing under unfavourable environmental conditions including low capillarity, salt spray, and dryness, water holding capacity, high light intensity, extreme temperature, sand burial, nutrient deficiency and negative action of violent winds (Gratani et al. 2009). These abiotic stress factors generate secondary osmotic and oxidative stresses, which then negatively influence and induce changes in the normal growth, development, and metabolism of the plant (Kròl et al. 2014). In spite of all these constraints, these environmental conditions promote the development and evolution of some particular species belonging to different botanical families. Many plant species have adapted on genetic, morphological and physiological levels, which helps them partially or completely to withstand these stress conditions (Gad et al. 2002). These established strategies are an important step that could explain the preservation and survival of these species in such extreme and hostile environmental conditions. To explore these strategies and in order to value plant resources native to saline biotopes, we have chosen four species belonging to two important families, Brassicaceae and Fabaceae.
The genus Matthiola W.T.Aiton (Brassicaceae) contains 50 species of annual and biannual herbs and perennial sub-shrubs, characterized by a silicle that features horns or swellings in its apical end (Sánchez et al. 2006). Matthiola tricuspidata (L.) W.T.Aiton is a small annual plant characterized by a sturdy stem softly drooping, sinuous leaves, large flowers and a potential oilseed (Demiral 2003); this species is widespread on the sand and rocks of the coast (Quezel and Santa 1962). Cakile maritima Scop. is an annual, succulent and halophyte plant (Ksouri et al. 2007), growing in sandy habitats along the North Atlantic Ocean, the Mediterranean Sea coasts, the Canary Islands and in southwestern Asia (Clausing et al. 2000). It is usually positioned in front of the pioneer dune and is fixed by its developed roots and it fleshy and pinnatifid leaves (Parisod and Baudière 2006). This plant has an ecological value as its roots bind dunes, potential economic use as its seeds contain oil (40% of dry weight), and a therapeutic value (Ciccarelli et al. 2010).
Among the Fabaceae two species have also been selected: Ononis variegata L. and Lotus creticus L. The genus Ononis L. comprises about 75 species occurring in Europe, Central Asia, and Africa (Wollenweber et al. 2003), being generally perennial herbs and shrubs (Wdowiak-Wróbel et al. 2017). Many species of the Ononis genus carry glandular trichomes that are more or less viscent, and contain flavonoids in several cases (Wollenweber et al. 2003). The genus Lotus L. comprises between 120 to 130 species of perennial and annual native plants, distributed mainly in the Mediterranean region (Sandral et al. 2010). In this genus, L. creticus is considered an important perennial species due to its ability to stabilize dunes (Sandral et al. 2010).
Plants living under hostile conditions synthesize higher amounts of protecting specialized metabolites such as phenolic compounds with free-radical quenching activity which can be primarily related to the substitutions on the aromatic ring and the structure of the side chain (Abideen et al. 2015). These compounds are involved in various plants physiological and developmental processes such as growth regulation, pigmentation, reproduction, and resistance to pathogens. They protect plants against adverse factors including drought, UV radiations, infection and physical damage (Slatnar et al. 2016). Various biotic and abiotic factors generate oxidative stress that can be very harmful to cells (Xiao et al. 2014). In such situations, antioxidant molecules intervene and decrease the deleterious effect of free radicals (Apel and Hirt 2004). Thus, we evaluated the contents of total phenols, total flavonoids (one of the largest groups of plant phenols) and anthocyanins (a sub class of flavonoids) in the leaves of the four selected plants growing on the coastal dunes of Zemmouri El Bahri, Algeria. Moreover, major photosynthetic pigments and certain morphological parameters were also determined as they are usually affected by drought and other environmental conditions. In this paper, we compared morphological and physiological traits of Matthiola tricuspidata, Cakile maritima, Lotus creticus and Ononis variegata with the aim of exploring their possible adaptive mechanisms to environmental stress.
Materials and methods
Study site and plants materials
The study was conducted in the coastal dunes of East-Algiers (Zemmouri El Bahri), located between 3°32', 3°38' E and 36°48', 36°50' N geographical coordinates. The area is characterized by a Mediterranean climate, with an annual mean rainfall of 579 mm. The mean air temperature of the coldest months reaches 9.3 °C whereas the mean maximum air temperature of the warmest month goes up to 28.9 °C, with a drought period spread over five months, a subhumid bioclimatic zone and a variant hot winter (Hanifi et al. 2007). The species M. tricuspidata, C. maritima, L. creticus and O. variegata were collected at the floral stage during April and May from the study station (sand dunes of Zemmouri el Bahri). The leaves were freshly harvested, washed, dried, finely ground and then stored in a dry dark place.
Morphological parameters of the leaf
The morphological leaf traits were determined by the method described byCiccarelli et al. (2010). The leaves from three individual plants per selected species were collected, and weighed to determine the fresh weight (FW). Leaf projected area (LA) was obtained with a Canon Scan using a Meserium Software area and expressed in cm2. Plant material was dried at 70 °C to constant weight (dry mass, DM).
Leaf dry matter content (LDMC) was calculated as the ratio of leaf dry mass (DM) to fresh mass (FM), expressed in mg g-1 (Li et al. 2005):
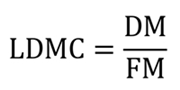
Leaf mass per area (LMA) was expressed as the ratio of dry mass (DM) to leaf area (LA), expressed in mg cm-2 (Ogaya and Penuelas 2007):
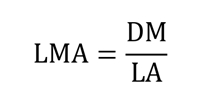
Succulence index(SI) is the ratio of the difference between the dry mass (DM) and fresh mass (FM) to the leaf area (LA), expressed in mg cm-2 (Gratani et al. 2009):
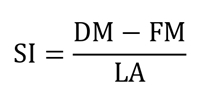
Where DM corresponds to: dry mass, FM: fresh mass, LA: leaf area.
Physiological parameters
Leaf relative water content (RWC) was evaluated using the equation introduced byLadigues (1975):
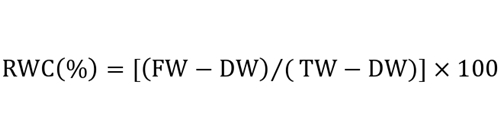
Where FW corresponds to: fresh weight, DW to: dry weight and TW to: full turgor weight, measured after soaking the leaves in water for 24 hours at room temperature and in the presence of light. The dry weight was estimated after drying the leaves at 60 °C for 48 hours (Clarck and Mac-Caig 1982).
To extract the photosynthetic pigments chlorophyll a (Chl a) chlorophyll b (Chl b), total chlorophylls (Chl a+b) and total carotenoids (Car), 0.05 g of fresh leaf samples were ground in 5 mL of acetone solution (80%). The homogenate was centrifuged at 3000×g for 10 min at 4 °C. The supernatant was recuperated and the absorbance was measured at 647 nm, 663 nm and 470 nm for Chl a, Chl b and Car, respectively. The amounts of pigments present in the extract were calculated using the equation ofLichtenthaler (1987):

Biochemical parameters
Sample extract was obtained by stirring 1 g of dry leaf powder with 7 mL of pure methanol and 3 mL deionized water for 30 min. The extracts were then kept for 48 hours at 4 °C, filtered through N°1 Watman filter paper, evaporated under vacuum to dryness and then stored at 4°C until analysis (Ksouri et al. 2007).
Total phenolic contents were determined using the Folin-Ciocalteu reagent, according to the method ofSingleton and Rossi (1965). A volume of 0.025 mL of sample extract was added to 3.975 mL of distilled water before adding 0.25 mL of the Folin-Ciocalteu reagent. The mixture had the ability to react for 3 minutes before 0.75 mL of 20% Na2CO3 was added. After incubation for 40 minutes at 40 °C, the absorbance was measured at 760 nm. There was a sequence of three assays, and the phenolic contents were expressed as Gallic acid equivalent in milligrams per gram of dry weight (mg GAE g-1 DW).
Total flavonoids were measured using a colorimetric aluminum chloride (AlCl3) assay (Lamaison and Carnet 1990,Chang et al. 2002). A volume of 1.5 mL of each extract was mixed with 1.5 mL of 2% AlCl3 6H2O. The preparation was shaken and the absorbance was read at 440 nm after 10 minutes at room temperature. The assay was repeated three times and the total flavonoid contents were expressed as rutin equivalent in milligrams per gram of dry weight (mg ERu g-1 DW).
The anthocyanin content was determined according to the procedure described byGould et al (2002) as follows; anthocyanins were extracted by the addition of 2 mL of acidified methanol 1% HCl (V/V) to 50 mg of dry leaves. The supernatant aliquot was retrieved and the absorbance was measured at 530 nm. Total anthocyanin was calculated by endoresing the following formula:

The anthocyanin content was expressed in mg of Cyanidin-3-glucoside equivalent per gram of dry weight.
Statistical analysis
The statistical study was performed with Excel and Statistica v.8 (StatSoft, Inc., Tulsa, OK). Results were analysed by one-way ANOVA, followed by Tukey test to compare means. The differences were held significant if P < 0.05. All experiments were carried in triplicates and expressed ± SD (n = 3).
Results
Plant morphological study
There were significant differences among the species for all morphological traits studied. Leaf succulence index (SI) shows different values in the four species (Tab. 1). Cakile maritima had the largest leaf area and the highest succulence index (12.51 ± 0.03 cm-2, 131.37 ± 0.83 mg cm-2, respectively), though both species of Brassicaceae showed high leaf area and a high succulence index. Fabaceae family species had narrow leaves, while L. creticus leaves exhibited the lowest leaf succulence (22.86 ± 0.56 mg cm-2). Leaf mass area (LMA) was also evaluated, this parameter varied from 6.55 ± 0.45 mg cm-2 (L. creticus) to 8.52 ± 0.74 mg cm-2 (C. maritima). Leaf dry matter content (LDMC) indicates that Fabaceae species had the highest values with 219.51 mg g-1, 147.64 mg g-1 for L. creticus and O. variegata, respectively. Among the Brassicaceae family species, C. maritima leaves showed the lowest LDMC (58.52 ± 0.53 mg g-1). Leaf area (LA) of C. maritima was significantly higher, but the lowest LDMC was found there. On the other hand, the leaves of L. creticus had the smallest foliar surfaces but highest dry matter accumulation (Tab. 1).
Plant physiological responses
The relative water content (RWC) was significantly higher in C. maritima (77.14%) than in the other species, in which it ranged from 63.10 to 65.15% (Fig. 1).

The variations of photosynthetic pigments are shown inFig. 2A. Among the four species, M. tricuspidata and L. creticus showed the highest levels of chlorophyll a (1.02 ± 0.04 mg g-1 FW and 1.11 ± 0.09 mg g-1 FW), total chlorophylls (1.39 ± 0.1 mg g-1 FW and 1.34 ± 0 .12 mg g-1 FW, respectively) and carotenoids content (0.41 ± 0.06 mg g-1 FW, 0.47 ± 0.08 mg g-1 FW), while the lowest contents of chlorophyll a, total chlorophylls and carotenoids were obtained for C. maritima. The content of Chl b was similar for all the species and the Chl a/b ratio varied from 2.82 (M. tricuspidata) to 2.33 (L. creticus). The chlorophyll/carotenoid ratio was in the range of 3.42 to 2.78 (Fig. 2B).

Plant biochemical responses
The results of total phenolic compounds and total flavonoids in plant extracts are presented inTab. 2. The results obtained show that the content of total phenolic compounds varied between different species; M. tricuspidata had the highest total phenolic compounds with 72.72 ± 0.45 mg GAE g-1 DW, and the lowest were found in C. maritima (24.17 ± 0.25 mg GAE g-1 DW). The quantities of total phenolic were 40.73 ± 0.77 mg GAE g-1 DW and 50.01 ± 0.37 mg GAE g-1 DW for L. creticus and O. variegata, respectively. Flavonoid content also varied among the species; M. tricuspidata showed the highest content (7.36 ± 0.09 mg ERu g-1 DW), and the lowest was found in C. maritima (1.93 ± 0.02 mg ERu g-1 DW) which is 4 times lower than that of M. tricuspidata. The values recorded for Fabaceae species were 5.86 ± 0.21 mg ERu g-1 DW and 3.7 ± 0.08 mg ERu g-1 DW for L. creticus and O. variegata, respectively.
The results of anthocyanin contents of species present some notable differences (Fig. 3). Leaves of M.tricuspidata displayed the highest levels of anthocyanins with 0.54 ± 0.06 mg g-1 DW, compared to other species. The values of those compounds were 0.36 ± 0.05 mg g-1 DW for O. variegata, 0.29 ± 0.05 mg g-1 DW for L. creticus and 0.25 ± 0.02 mg g-1 DW for C. maritima.

Discussion
The results of the study show that Matthiola tricuspidata, Cakile maritima, Lotus creticus and Ononis variegata living on the sand dunes have different morphological, physiological and biochemical adaptive strategies to survive in the hostile conditions of the coastal dunes. Drought, salinity and UV intensity affect the photosynthesis, growth and development of species (Shah et al. 2017). Relative water content is probably the most appropriate measure of plant water status in terms of the physiological consequence of cellular water deficit (Tátrai et al. 2016). Cakile maritima showed the highest relative water content (RWC), a large leaf area (LA), leaf mass area (LMA) and succulence index (IS), but its Chl a, Chl b and total chlorophyll contents were the lowest. The high LMA represents a morphological character of Mediterranean species that plays a protecting role for plants exposed to drought stress (Gratani et al. 2009). The values of leaf mass area (LMA) and index of succulence (IS) are similar to those reported in some species of the Mediterranean coastal dunes (Gratani et al. 2009). Cakile maritima is a halophyte and succulent plant, which is naturally tolerant under conditions of salinity and drought (Ksouri et al. 2007), with very developed taproots allowing it maximum water absorption, and fleshy leaves, ensuring the accumulation of large tissue water reserves (Parisod and Baudière 2006). The survival of plants in circumstances of water deficiency is dependent on their osmotic adjustment (Baroowa and Gogoi 2015). Different studies have indicated that the drought stress decreases Chl a, Chl b and the total chlorophyll content of the various crops (Hailemichael et al. 2016). In this regard, C. maritima could be classified as one of the "poikilochlorophyllous" species (Challabathula et al. 2018) that reduce their chlorophyll concentration to absorb less light energy and thus have less excess electrochemical energy, and therefore their needs for antioxidant defense are less important. The content of phenolic compounds in C. maritima is close to that reported byMeot-Duros et al. (2008): 22.24 mg EAG g-1 DW andKsouri et al. (2007): 42.84 ± 7.67 mg EAG g-1 DW. The total content of flavonoids in plants from the Brassicaceae family ranged from 0.72 EC.g1 DW to 15.38 EC.g-1 DW (Grigore and Opricã 2015), close to the values reported in this study.
The flavonoid content found in Lepidium crassifolium (2.37 EC.g-1 DW) was similar to that in C. maritima (1.93 mg ERu g-1 DW).Ivan and Opricã (2013) also reported total flavonoid levels comparable to those in our results. Cakile maritima adopts a "drought avoidance" strategy to resist drought, which is defined as the ability of the plant to maintain a relatively high water content in tissues despite reduced water content in the soil (Omprakash et al. 2017). In comparison with C. maritima, M. tricuspidata had a lower relative water content and higher content of Chl a and Car. The levels of total phenols, total flavonoids, and anthocyanins were the highest of all the species. Based on those results, M. tricuspidata can be classified as one of the "homoiochlorophyllous" species that maintain their chlorophyll pool relatively high under high salinity and light intensity (Challabathula el al. 2018). As a large part of the light energy captured and converted into electrochemical energy at the reaction centers would then be in excess, these species need to have efficient protective mechanisms against the free radicals formed. The carotenoids (xanthophylls) are necessary for photoprotection of photosynthesis and they play an important role as a precursor in the signaling during plant growth under abiotic/biotic stress (Ola and Abd El-Maboud 2013). This could explain the high levels of antioxidant molecules such as carotenoids, total flavonoids and anthocyanins that we have recorded in M. tricuspidata. The chlorophyll content results reported in this study were similar to those found in plants growing in coastal dunes (Frosi et al. 2017). Many studies have shown that the concentrations of specialized metabolites in plant exposed to drought are higher than in those grown under well-watered conditions (Al-Gabbiesh et al. 2015).
The results of total phenol contents of species are in accordance with studies on several halophytes (Ksouri et al. 2007,Ivan and Opricã 2013). An increase of content in phenolic compounds in the different tissues under the effect of salinity has been recorded in a certain number of plants (Ksouri et al. 2007,Abideen et al. 2015). Plants react to various environmental factors by increasing the production of polyphenols, particularly flavonoids (Anket et al. 2019). These molecules have important properties such as the protection of tissues against the harmful effects of UV radiations and are an important antioxidant activity (Xiao et al. 2014).
The two species of the Fabaceae family have different morphological strategies; L. creticus and O. variegata have smaller leaf areas (LA), lower leaf mass area (LA), and succulence index (SI). The two species show a combination of high leaf dry matter content (LDMC) and low leaf mass area (LMA). The species with high LDMC and low LMA show a slow growth rate, with long leaf lifespan and more efficient conservation of nutrients (Saura-Mas and Lloret 2007). The reduced leaf area is a strategy to conserve water and control excessive transpiration (Melo Júnior and Boeger 2016). The species with low LMA (i.e. high specific area, SLA), is characterized by high photosynthetic capacity per unit mass (Gratani et al. 2009). Since both L. creticus and O. variegata displayed high contents of chlorophylls, carotenoids, total phenols and flavonoids, they could also be classified as "homoiochlorophyllous" species. The results of total phenolic and flavonoid contents were similar to those reported bySayari et al. (2016).
Thus, M. tricuspidata, L. creticus and O.variegata use the "drought tolerance" strategy to resist water deficit, which involves maintaining cell turgor by osmotic adjustment, the elevation of protoplasmic resistance (Supratim et al. 2016), the extension of antioxidant capacity, and development of desiccation tolerance (Osmolovskaya et al. 2018).
Conclusion
This paper shown the various strategies adopted by the four species under unfavorable environmental conditions present in coastal dunes. On the morphological and physiological level, C. maritima has a high water potential (high RWC) with a succulent character, which is an adaptive trait that comprises a large leaf area and water reserve. In comparison, M. tricuspidata, L. creticus and O. variegata show lower relative water content, have relatively high chlorophyll contents and fight the consequent higher oxidative damage by a greater accumulation of antioxidant molecules such as phenolic compounds and carotenoids. In addition, L. creticus and O. variegata showed reduced leaf area and a high leaf dry matter content, i.e. leaf morphological adaptations enabling protection against water deficit and limitation of water losses.