
Introduction
The family Combretaceae is an advanced flowering plant with a worldwide tropical distribution (Hutchinson 1969). The family has a different number of genera, ranging from 16 (Rendle 1956) to 20 (Mabberley 2017). Cronquist (1981) recognized 400 species, while Willis (1966) recognized 600 species within this family. These variations in the number of genera and species are due to a lot of synonyms and the either clumping or splitting of genera (Scott 1979). Classification of Combretaceae has been faced with many opinions since Exell (1954) as the family was divided into two subfamilies, Strephonematoideae with the monotypic genus Strephonema and Combretoideae with the rest of the genera. More recent treatments by Exell and Stace (1966) and Stace (2007), divided the family into two tribes, Laguncularieae and Combreteae. The latter tribe was subdivided into three subtribes: Combretinae, Pteleopsidinae and Terminaliinae. Terminalia is the second largest genus after the genus Combretum under the subtribe Combritenae, tribe Combreteae, family Combretaceae, suborder Myrtineae, order Myrtales, (Stace 2002, Heywood et al . 2007). Species under this genus are shrubs, trees, and creepers distributed in tropical and subtropical regions, especially in Africa and savannas (Arundhati et al. 2020). Many of its species have high economic value as they are used widely in commercial applications and Chinese traditional medicine (Zhang et al. 2019). The genus name came from the word terminal in Latin, as their leaves are crowded at the tops of the shoots. Recognition of the species under the genus Terminalia was according to fruit and leaf morphological characters (Chakrabarty et al. 2019, Rayane et al. 2022). The species under this genus differ greatly in their internal and external features in addition to their chromosome numbers (Stace 1965, Ohri 1996). Wickens (1973) and Fyhrquist (2007) found that bark, leaves, and fruit morphology can be used as diagnostic features in the identification of Terminalia species. Since species under the genus Terminalia are traditionally used in the treatment of some illnesses, precise identification and recognition of the closely related species need clarification.
Flowers in the genus Terminalia are grouped in terminal simple racemes with duplicates of five anthers alternating with five petals. Despite the numerous stamens, only a few pollen morphological studies have dealt with taxonomical issues of the genus, e.g. Erdtman (1966), Patel et al . (1984), El-Ghazali et al. (1998), Krachai and Pornpongrungrueng (2015). Other palynological articles were mostly focused on the family level. The taxonomic disagreement concerning the species in the genus Terminalia, compounded by the challenge of accurately identifying certain species based only on macro-morphological characteristics, is the reason for this investigation. The aims of this work are to investigate the variation in the pollen pheno-characters of Terminalia species, and to classify the different species under this genus with respect to their pollen characters. Therefore, the subject of this investigation was the pollen grain classification of eleven Terminalia cultivated species, planted in Cairo parks, streets, and botanical gardens as ornamental trees.
Material and methods
Mature unopened flowers of eleven species were collected from Egyptian botanical gardens and streets during the flowering season (Tab. 1). The flowers were opened carefully using two needles under a stereo-microscope to release the anthers. The obtained anthers were smeared onto cleaned glass slides with a few drops of glycerol, covered, and sealed with paraffin wax. In order to prevent the harmomegathic effects − which occur when chemicals are exposed to aperturate pollen grains during the acetolysis process and the grains fold in response to changes in humidity (Taia 2022), non-acetolysed pollen grains were examined and then measured and photographed using a Motic (B-150D) light microscope (LM) fitted with a USB digital-video camera and computer software with 10× and 40× objective lenses.
All the measurements were based on 20 to 30 pollen grains. A calibrated oculometer was used to obtain the minimum and maximum measures and calculate the mean and standard deviation of each parameter and the apocolpium index (polar area index), which was calculated as the ratio of the mean distance between the apices of two colpi in both the main and sub apertures to its equatorial diameter. Values are expressed in micrometers.
For scanning electron microscope (SEM) investigation, non-acetolysed pollen grains were sputtered onto double sticks fixed in cleaned, aluminum labeled stubs, then coated with 20 nm gold in a Polaron JFC-1100 coating unit, finally examined and photographed using JEOL-JSM.I T200 Series Scanning Electron Microscope made available in the electron microscope unit, Faculty of Science, Alexandria University, Egypt. The terminology used for the descriptions is that of Punt et al. (2007).
Tab. 1. The localities of collected specimens, geographic coordinates, and the date of collection of the investigated Terminalia species, kept at Cairo University Herbarium (CAI).
Fifteen pollen morphological characters have been subjected to cluster analyses using PAST 3 programs V.3. designed by Hammer (1999-2013).
To generate a key, the data obtained from the description of pollen characters were subjected to automated key generation using version 4.12 of the DELTA suite of programs (Dallwitz et al. 2000).
Results
General description of Terminalia pollen grains
For the accurate identification of members of the Terminalia species, the morphological characteristics of pollen − such as size, shape, aperture conditions, and exine ornamentation − are crucial. The morphological properties of pollen, both quantitative and qualitative, derived by LM and SEM analysis are compiled in Tab. 2 and Fig. 1 and Fig. 2. The examined species' pollen grains are eurypalynous, meaning they vary greatly in character, shed in monads, apolar or isopolar and radially symmetric. These species are classified as either monomorphic or dimorphic. T. arjuna, T. bentzoe, T. catappa, T. mantaly, and T. muelleri are examples of monomorphic plants; on the other hand, the dimorphic species were represented by T. bellirica, T. brownii, T. chebula, T. laxiflora, T. myriocarpa and T. sericea; indicating that they have two distinct pollen shapes, sizes, apertures, and even exine ornamentation within the pollen grains that are gathered from the same flower.
Tab. 2. Pollen morphological data of 11 Terminalia species from Cairo, Egypt. AI − apocolpium index, Amb − the outline of pollen grain seen in polar view, Cir − circular, Di – dimorphic, E− equatorial axis diameter (μm), Eth − exine thickness (μm), EO − exine ornamentation, Gr − group, Hlob − hexalobate, M – mean, Mono – monomorphic, Min–Max – minimum and maximum values, No − number, P − polar axis length (μm), P/E − polar axis/equatorial axis, Pr − prolate, RG − rugate granulate, RP − rugate perforate, Sca − scabrate, SD – standard deviation, Sh − shape, SP − sub prolate, Sph − spheroidal, St − striate, StR − striate rugate, Tr − triangular.
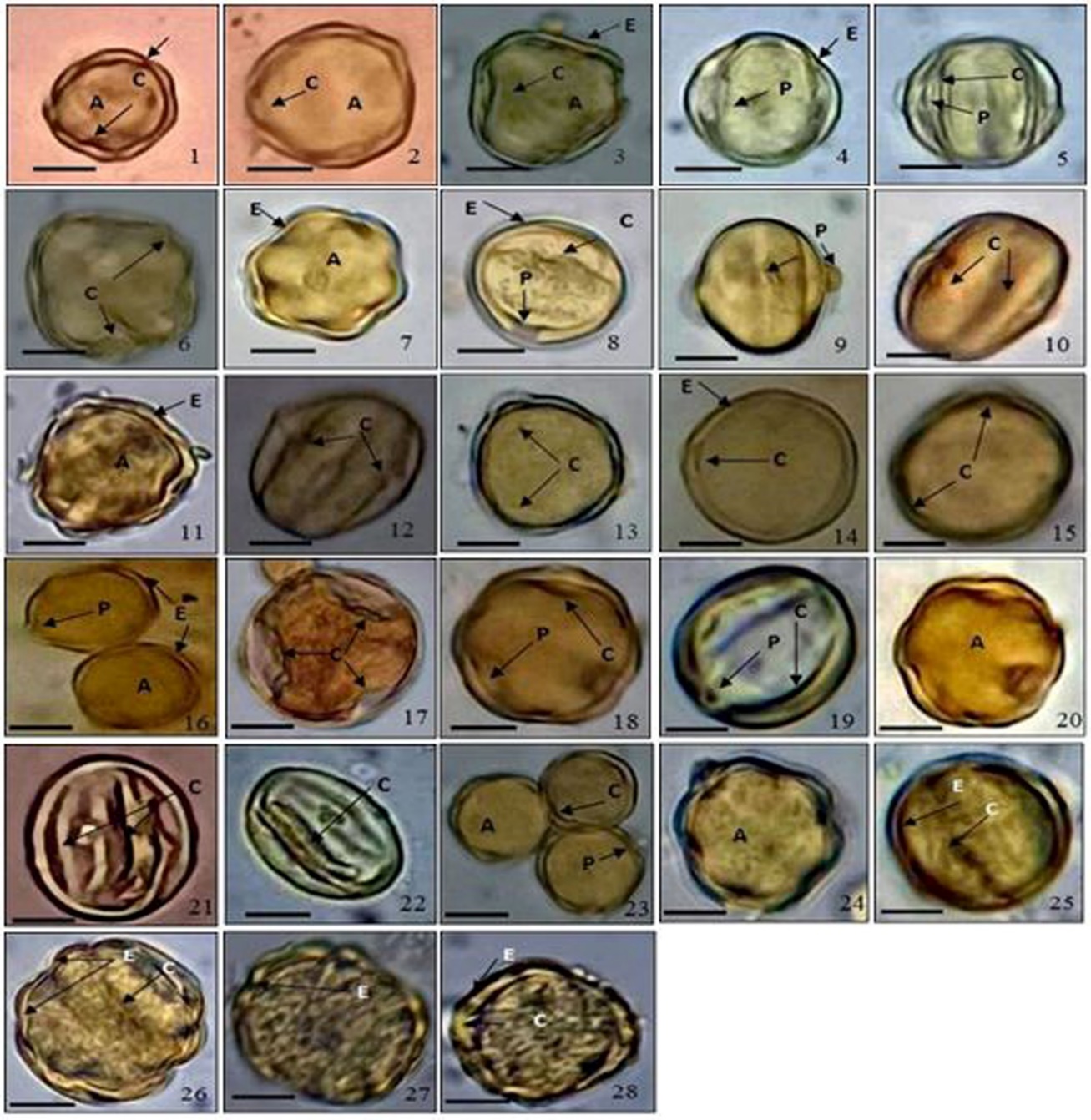
Fig. 1. Light microscopy micrographs of the studied Terminalia species. (1-2) T. arjuna, (3-6) T. bellirica, (7-8) T. bentzoe, (9-10) T. brownii, (11- 12) T. catappa, (13- 16) T. chebula, (17-19) T. laxiflora, (20) T. mantaly, (21-22) T. muelleri, (23-25) T. myriocarpa, (26-28) T. sericea. (A − amb, C − colpus, E − exine, P − pore, bar = 10 µm).
Shape and size
In their polar view, the pollen grains were triangular, hexa-lobed or circular (Fig. 1 and Fig. 2). In the equatorial view, they were spheroidal ( T. bellirica, T. bentzoe, T. chebula, T. mantaly and T. sericea), subprolate ( T. arjuna, T. bellirica, T. brownii, T. chebula, T. laxiflora, T. muelleri, T. myriocarpa and T. sericea) and prolate ( T. brownii, T. catappa, T. laxiflora and T. myriocarpa). T. catappa had the largest pollen diameter on the polar axis (P) at 34.6 μm, while T. sericea had the smallest (18.6 μm). Similarly, T. muelleri and T. myriocarpa had the biggest pollen diameter on the equatorial axis (E) (30.8 μm), whereas T. sericea had the smallest diameter (17.2 μm). Based on the ratio between the lengths of the polar axis (an imaginary straight line connecting the two poles) to the equatorial diameter (P/E ratio), pollen sizes and shape variations were examined; T. brownii had the highest P/E value (1.26), while T. bellirica, T. chebula, and T. sericea had the lowest values (0.1).

Fig. 2. Scanning electron microscope microphotographs showing the different pollen characters within the studied Terminalia species. (29-31) T. arjuna, (32-34) T. bellirica, (35) T. bentzoe, (36-37) T. brownii, (38-39) T. catappa, (40-41) T. chebula, (42-46) T. laxiflora, (47-49) T. mantaly, (50-52) T. myriocarpa. (a − amb, c − colpus, e − exine, p − pore)
Aperture conditions
Apertures are the first characteristic to take into account when identifying pollen. In T. arjuna, T. brownii, and T. catappa, the apertures are iso-colpate; in the other investigated species, the apertures are hetero-colpate; they have two or more types of colpi, one of which varies in length and/or in whether endoapertures are present. The six apertures seen on the pollen grains of the Terminalia species under study are composed of three primary, long apertures that alternate with three smaller, possibly distinct, but somewhat shorter apertures.
Pollens of the species T. arjuna are hexa-colpate in which the main apertures and the sub-apertures are of the same type. The pollen grains of the species T. bellirica, T. bentzoe, T. chebula, T. mantaly and T. myriocarpa are hexa-colpate in which the main apertures are colpate and the sub-apertures are colporate. Also, hexa-colpate pollen grains are observed in T. laxiflora, T. muelleri and T. sericea in which the main apertures are colpate and the sub-apertures are colporoidate. In T. brownii and T. catappa the pollen grains are hexa-colporate in which the main apertures and the sub-apertures are of the same type.
The colpi are not connected at the poles and the apocolpium indices varied between the studied species from 0.14 in T. brownii to 0.42 in T. sericea. The aperture membranes are either psilate or granulate. In the case of composite apertures, the pores are either small and in the same colpi direction (lolongate) or oval shaped covered by granulated exinous layer (colporoidate). According to the state of the pollen grains, momo-or di- morphic and aperture types the studied species are grouped mainly in two main groups and four subgroups (Tab. 2).
Exine ornamentation
Exine sculpturing is the most distinguishing character of the species; in this study, the species have some variation in exine ornamentation; the exine is generally tectate, scabrate in T. bentzoe, and T. chebula; rugate in T. laxiflora; rugate perforate in T. catappa, T. myriocarpa and T. sericea with thin ectexine and thick endexine and intine layers as observed by light microscope investigation. Striate ornamentation is observed only in T. brownii while striate-rugate ornamentation is recorded in T. arjuna, T. bellirica, T. mantaly and T. muelleri (Tab. 2). Exine thickness ranges from the minimum (2.0 μm) in T. bellirica to the maximum (2.6 μm) in T. arjuna, T. chebula and T. muelleri (Fig. 1).
Clustering analysis
The phenogram resulting from the cluster analysis of fifteen palynological characters (On-line Suppl. Tab. 1) after coding according to their states (On-line Suppl. Tab. 2) divided the studied taxa into three groups at similarity distance 0.936. Group 1 has one species only, T. sericea (11). Group 2 holds eight species, subdivided into two subgroups at similarity distance 0.952, 2A with three species; T. brownii (4), T. laxiflora (7), T. myriocarpa (10); and subgroup 2B with five species; T. catappa (5), T. mantaly (8), T. muelleri (9), T. arjuna (1), T. bellerica (2). Group 3 includes two species, T. bentzoe (3) and T. chebula (6) (Fig. 3).
Computer-generated key
1. Pollen shape subprolate or prolate …………………………………………… 2
Pollen shape subprolate or spheroidal ……………………………………….. 3
Pollen shape prolate ……………………………………………….. T. catappa
Pollen shape spheroidal ……………………………………………………… 4
Pollen shape subprolate ……………………………………………………… 5
2. Amb shape triangular or hexalobate; Exine ornamentation rugate-granulate ………………………………………...………………………..…. T. laxiflora
Amb shape triangular; Exine ornamentation striate ……………….. T. brownii
Amb shape circular; Exine ornamentation rugate-perforate ….... T. myriocarpa
3. Amb shape circular or hexalobate; Exine ornamentation rugate-perforate …………………………………………………………………..… T. sericea
Amb shape triangular; Exine ornamentation striate-rugate ………... T. bellirica
Amb shape circular; Exine ornamentation scabrate …………….…. T. chebula
4. Exine ornamentation scabrate …………………………………...…. T. bentzoe
Exine ornamentation striate rugate …………………………….….. T. mantaly
5. Amb shape circular or hexalobate; Sub aperture type colpate …....... T. arjuna
Amb shape hexalobate; Sub aperture type colporoidate ……….… T. muelleri
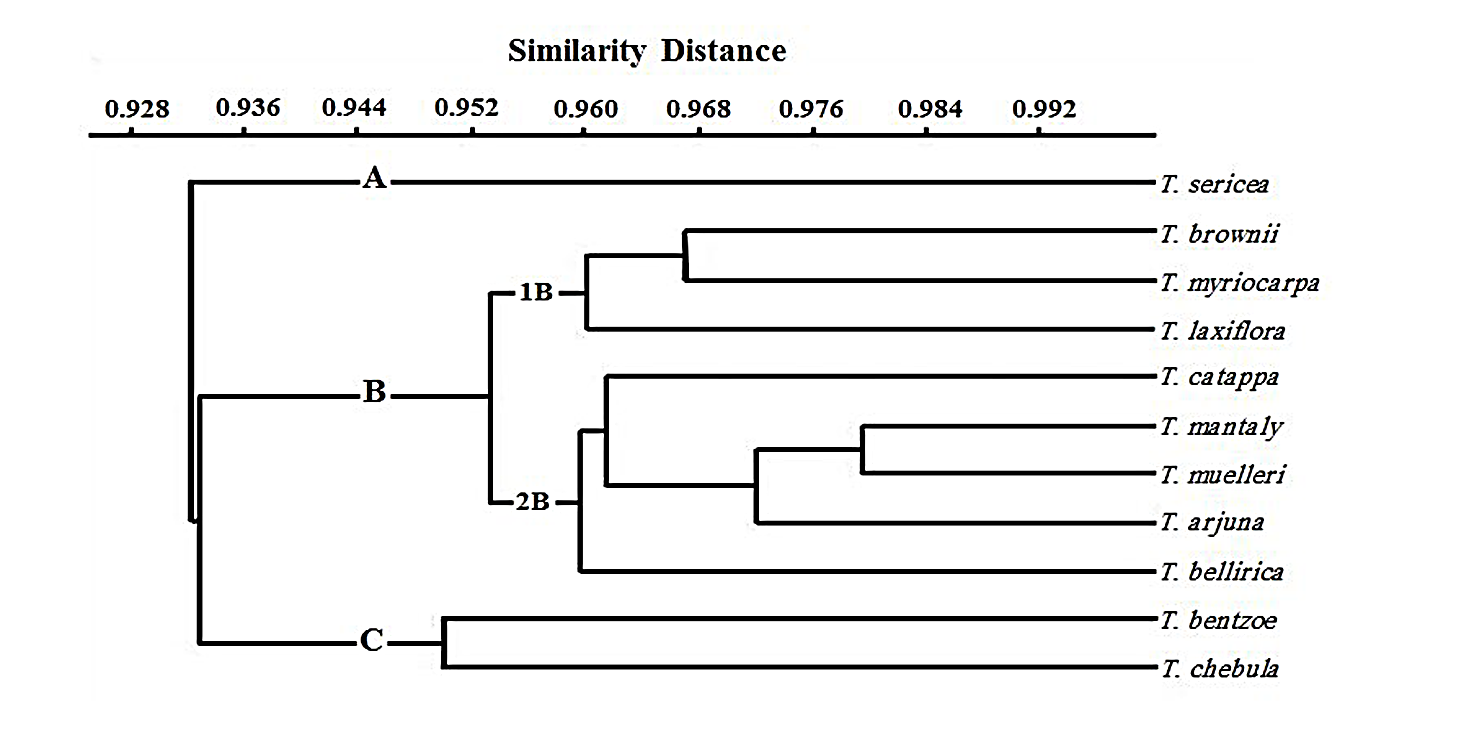
Fig. 3. Phenogram based on 15 pollen characters showing the possible relationships among the studied Terminalia species using percent similarity distances.
Discussion
Morpho-palynological studies have proved to be of great importance for plant identification and taxonomic classification. In this study, different palynological features such as variations in pollen characters in the same flower (morphism), polar and equatorial outline and their ratio (P/E), type of aperture, presence or absence of pores, beside exine thickness and ornamentation were investigated in eleven Terminalia (Combretaceae) species belonging to 7 sections from Egypt using LM and SEM. Based on palynological features, a taxonomic key was created for simple identification and species delimitations.
Mignot et al. (1994) pointed to the importance of aperture type in angiosperm recognition at any taxonomic level, even down to the intra individual level. Based on the results, Terminalia species may be divided into two main groups: monomorphic and dimorphic. Each group has a variety of pollen shapes, ranging from spheroidal to prolate, with many shapes seen in the pollen sampled from the same flower. Pollen hydration status and environmental conditions may contribute to heterogeneity in pollen shape and size within the same taxon (Grant-Jacob et al. 2022, Taia et al. 2023). El Ghazali (2022) noted differences in the sizes and forms of pollen among the same taxa within the family Combretaceae. Despite this, he defined their pollens as stenopalynous, meaning that their characteristics are either stable or slightly changed. According to this investigation, the Terminalia under study exhibit significant variety in their pollen morphological characteristics, and as a result, they can be classified as eurypalynous.
The highest PA length is seen in T. myriocarpa (27.8-34.5 µm), whereas T. sericea has the smallest PA length (18.6-23.2 µm). El Ghazali et al. (1998) have documented these differences in pollen size and shape. Given that multiple shape types have been observed in a single flower, shape class is determined to have less diagnostic value within the taxa in the material under examination. Taia et al. (2023) have noted this investigation; as a result, it does not play a significant role in the taxonomic discrimination of the examined taxa, except in the case of T. sericea, which is grouped separately in the clustering analysis (group A), this separation being due to it having the smallest PA and ED lengths and the smallest main aperture and sub-aperture lengths. This result supports the classification of this species as it belongs to section Psidioides Exell.
Four monomorphic species ( T. catappa, T. mantaly, T. muelleri, and T. arjuna) were gathered by the mono-morphic character within the same taxa and placed in subgroup 2B in close proximity to the dimorphic taxa ( T. bellerica). This grouping strengthens the taxonomic status of T. catappa, T. mantaly, T. muelleri as they belong to the same section Eucatappa Engl. & Diels. Despite T. arjuna being a separate section Pentaptera (Roxb.) Engl. & Diels, it is grouped with the former three species according to the mono-morphic pollen character.
According to El Ghazali and Krzywinski (1989), the apertures can be classified as iso- or hetero-colpate, meaning that they have two or more types of colpi, with one type differing in length and/or having endoapertures or not. This previously acknowledged observation was discovered in Rosa sericea (Ullah et al. 2022). It was also discovered in Fagonia species (Taia et al. 2021). El-Ghazali et al. (1998) noted the heterocolpate pollen grains within the taxa under study. This study noted variations in the length and width of the colpi as well as the existence and dimensions of endo-apertures; this result is in line with previous studies mentioned.
Since T. brownii, T. laxiflora, and T. myriocarpa have triangular or circular amb, the triangular amb gathered them in subgroup 1B. Amb shape was determined to be a valuable character in the building of the identification key. This result is partially in agreement with the taxonomy of T. brownii, T. laxiflora as they belong to the same section, Platycarpae Engl. & Diels, but does not agree with the classification of T. myriocarpa as a separate section Myriocarpae. A triangular amb is also present in T. bellirica, although it is distinct from the preceding three species in terms of other pollen characteristics and is located in subgroup 2B, albeit remaining closely linked to them. Certain species could be distinguished using the hexagonal, triangular, or circular amb, as the identification key illustrates. The two species; T. bellirica and T. bentzoe are belonging to section Billiricae Engl. & Diels, however, the study separated them in different groups; this separation is due to the difference between them in pollen state and amb shape.
Exine thickness ranged from 2.0 µm to 2.6 µm; however, there were significant differences in exine ornamentation. The rugate-perforate pattern was seen in T. myriocarpa and T. sericea, while the striate or striate-rugate pattern was seen in the majority of the investigated species. Only T. laxiflora has rugate-granulate ornamentation, but T. bentzoe and T. chebula have scabrate exine ornamentation; for this reason, the former two species were grouped together in group C although they belong to different sections; T. bentzoe sec. Belliricae Engl. & Diels, T. chebula sec. Myrobalanus (Gaertn.) Eichl. As a result, exine thickness varies very little among the taxa, whereas exine ornamentation, displayed by the key, is more helpful in identifying the taxa. This observation is consistent with every palynological study conducted since that of Erdtman (1966).
Conclusion
Based on the study's findings, three groups and two subgroups can be identified based on the unique characteristics of the pollen morphology of the Terminalia species under investigation. The pollen morphism, aperture homogeneity, amb form, and exine ornamentation are the most crucial characteristics in classifying the investigated taxa.