
UVOD - INTRODUCTION
Hrast kitnjak (Quercus petraea (Matt.) Lieblein) jedna je od ekonomski i ekološki vrlo važnih vrsta šumskog drveća i javlja se u oko 15% svih šuma u Bosni i Hercegovini (Matić i dr. 1971). Jedinke hrasta kitnjaka mogu biti vrlo dugovječne (preko 1 000 godina u nekim slučajevima) i mogu dostići visinu višu od 40 m te promjer od tri do četiri metra (Colin Prentice i Helmisaari 1991, Praciak i sur. 2013, Eaton i sur. 2016). Drvo hrasta kitnjaka i hrasta lužnjaka igralo je važnu ulogu u razvoju društava u Europi, osiguravajući drvo za ogrjev, plodove za ishranu stoke, koru za štavljenje i drvo za građevinarstvo (Eaton i sur. 2016).
Ducousso i Bordacs (2004) navode da su u prošlosti ljudi svojim aktivnostima značajno smanjili rasprostranjenost hrasta kitnjaka i hrasta lužnjaka, ali da se pokrivenost hrastovim šumama povećala od 19. stoljeća zahvaljujući organiziranom podizanju novih hrastovih šuma. U Europi postoji duga tradicija gospodarenja hrastovim šumama, koja se čini vrlo konzervativnom u pogledu genetskih resursa, ali je utjecaj različitih praksi podizanja novih šuma relativno nepoznat, navode Ducousso i Bordacs (2004), te ističu unošenje egzotičnih genotipova kroz umjetne nasade kao osnovnu prijetnju, koja je u prošlosti bila zanemarena. Ove populacije su u velikom riziku od nestanka, jer je broj jedinki s kojih se sakuplja sjeme mali, staništa su nestabilna i ljudski utjecaj je često značajan (Ducousso i Bordacs 2004).
Fenotipska varijacija odražava rezultat genetske varijacije i fenotipske plastičnosti kao odgovor na varijacije u okolišu. To je izraz prilagodbe biljaka na različite uvjete okoliša (Liu i sur. 2022), pa je fenotipska varijacija važna u istraživanju prilagodbe okoliša i evolucije biljaka (Edelar i sur. 2017). Proučavanje varijacija svojstava važno je za genetsko poboljšanje i očuvanje genetskih resursa hrasta kitnjaka. Ta istraživanja mogu pomoći u razumijevanju mehanizama reakcije, odnosno odgovora i obrazaca varijacija biljaka na okoliš, što je važno za prikupljanje, očuvanje i procjenu resursa biljnih germplazmi (Wang i sur. 2023). Varijacije prirodnih populacija hrasta kitnjaka kod nas i u okruženju nisu dovoljno istražene i hitno su potrebna daljnja istraživanja.
Postojeće studije o fenotipskoj (Bruschi i sur. 2003) i genetičkoj raznolikosti (Gömöry 2001, Ballian i sur. 2007, Ballian i Bogunić 2012, Ballian 2016, Bruschi i sur. 2003a, Rebrean i sur 2023, Tóth i sur. 2023), navode postojanje unutarpopulacijske i međupopulacijske varijabilnosti hrasta kitnjaka u Bosni i Hecegovini i različitim dijelovima Europe. Bruschi i sur. (2003a) navode postojanje diferencijacije populacije prema morfološkim i molekularnim biljezima, ali se obrasci diferencijacije morfoloških svojstava nisu poklapali sa mikrosatelitnom diferencijacijom, jer na molekularnoj razini nije bilo interakcija s okolinom. U pogledu fenotipske varijacije u morfološkim svojstvima lista i anatomiji hrasta kitnjaka Bruschi i sur. (2003b) utvrdili su velike varijacije unutar stabla zbog položaja grana, posebice u debljini i produktivnosti (40%). Za 19 od 32 varijable, varijacije među stablima bile su niže od varijacija unutar stabla, što su objasnili položajem grana (Bruschi i sur. 2003).
Informacije o fenotipskim varijacijama hrasta kitnjaka i njihovim odnosima s geoklimatskim faktorima na razini populacije ključne su za planiranje mjera genetskog očuvanja i unapređenja stanja populacija hrasta kitnjaka.
Naši glavni ciljevi bili su (1) kvantificirati fenotipsku varijaciju analiziranih svojstava listova; (2) odrediti udio fenotipske varijacije između i unutar populacija; i (3) ispitati obrazac varijacije populacija i njegovu povezanost s geoklimatskim faktorima. Rezultati ovoga istraživanja trebali bi pružiti osnovu za genetsko poboljšanje, očuvanje raznolikosti, uzgoj šuma i upravljanje resursima hrasta kitnjaka u Bosni i Hercegovini.
MATERIJAL I METODE - MATERIAL AND METHODS
Materijal - Material
Materijal je prikupljen sa po osam do deset stabala po populaciji u 24 prirodne populacije u Bosni i Hercegovini tijekom 2023. godine. Osnovni podaci o populacijama prikazane su u tablici 1.
Tablica 1. Geolokacije istraživanih prirodnih populacija Q. petraea
Table 1. Geolocations of investigated natural populations of Q. petraea
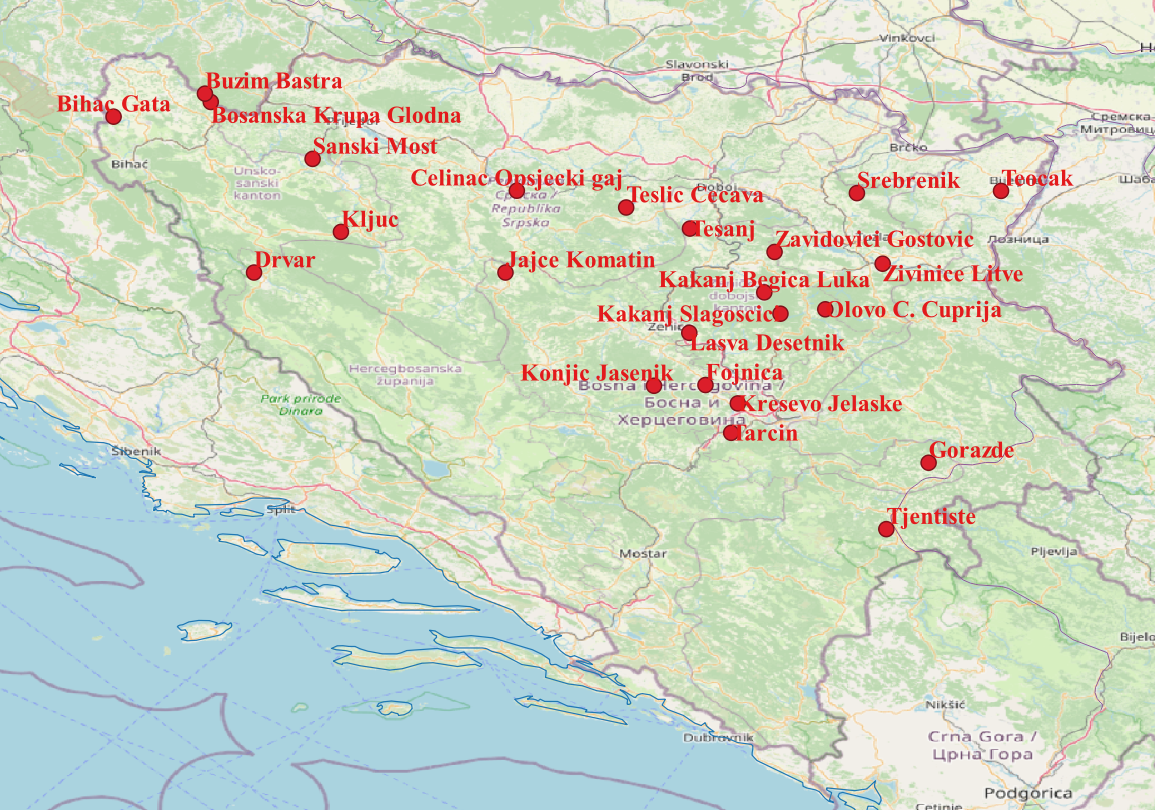
Slika 1. Geolokacije 24 uzorkovane populacije Quercus petraea
Figure 1. Geolocations of the 24 populations sampled of Quercus petraea
Zemljopisne lokacije uzorkovanih populacija određene su pomoću GPS-a, a sastoje se od zemljopisne širine, dužine i nadmorske visine. U istraživanju su korišteni bioklimatski podaci preuzeti sa WorldClim portala (https://www.worldclim.org/data/bioclim.html, pristup 10.05. 2022.). Nazivi klimatskih elemenata prikazani su u tablici 2.
Tablica 2. Lista bioklimatskih varijabli (WorldClim baza podataka)
Table 2. List of bioclimatic variables (WorldClim database)
Listovi hrasta kitnjaka sakupljeni su sa plodnih izbojaka (kratkorasta) i južno eksponiranog dijela krošnje adultnih stabala na osami ili stabala na rubovima šuma, koja su se u ranijim istraživanjima pokazala podobnim za morfometrijske analize, te dobri reprezenti istraživanih populacija (Trinajstić 1988, Franjić 1994, 1996 a, 1996b). Sakupljeno je po 30 listova po stablu. Listovi su zatim herbarizirani, osušeni te mjereni.
Morfometrijske analize - Morphometric analyses
Suhi listovi su mjereni digitalnim pomičnim mjerilom (u mm). Na svakom listu izmjereno je šest fenotipskih svojstava (Slika 2): duljina plojke (DPL), duljina peteljke (DPT), udaljenost najšireg dijela plojke od baze plojke sa desne strane (UNDP), najveća širina desne poluplojke (NSDP), najveća širina lijeve poluplojke (NSLP), širina usječenog dijela plojke ispod najveće širine sa lijeve strane (SUD). Izračunata je ukupna duljina lista (duljina plojke + duljina peteljke) (UD), ukupna širina plojke (širina lijeve poluplojke + širina desne poluplojke) (US), odnos širine naspram duljine lista (OSD) i odnos duljine peteljke naspram duljine plojke (OPP). Izvršeno je brojanje režnjeva sa desne strane (BR), te ocjenjivanje dlakavosti (DL) u četiri kategorije; 1 – nema, 2 – u pazuhu lista, 3 – na cijelom listu, 4 – uz središnji nerv) i ocjenjivanje oblika baze lista (OBL) (prema shemi, Slika 2).

Slika 2: Mjerene i ocjenjivane karakteristika lista: DPL- duljina plojke, DPT - duljina peteljke, UNDP - udaljenost najšireg dijela plojke od baze plojke sa desne strane, NSDP - najveća širina desne poluplojke, NSLP - najveća širina lijeve poluplojke, SUD - širina usječenog dijela plojke ispod najveće širine sa lijeve strane; OBL - oblik baze lista prema Fortini et al. (2015)
Figure 2: Measured and evaluated characteristics of the leaf: DPL - length of the leaf blade, DPT - length of the leaf petiole, UNDP - distance of the widest part of the blade from the base of the blade on the right side, NSDP – maximum width of the right half blade, NSLP - maximum width of the left half-blade, SUD- width of the incised part of the leaf below the maximum width on the left side; OBL - leaf base shape according to Fortini et al. (2015)
Statistička analiza - Statistical analysis
Deskriptivna statistika obuhvaćala je maksimalnu, minimalnu, prosječnu vrijednost, standardnu devijaciju (SD) i koeficijent varijacije (𝐶𝑉) kao relativna mjera varijabiliteta, koristeći sve podatke. Koeficijent varijacije izračunat je kao:
Koeficijent diferenciranja populacija (Vst) je izračunat kao:
gdje je -varijansa između populacija, a – varijansa unutar populacija (Wang i dr. 2023). i su dobijene primjenom ugniježđene analize varijance (ANOVA) (faktori stabla ugniježđeni unutar faktora populacije), prema sljedećem linearnom modelu:
gdje je: k-ta vrijednost na j-tom stablu u i-toj populaciji, u je ukupan prosjek, je slučajna (random) efektivna vrijednost u i-toj populaciji, je slučajna efektivna vrijednost j-tog stabla u i-toj populaciji, i je eksperimentalna greška ijk-te opservacijske vrijednosti, što je varijacija unutar stabala.
Testiranjem homogenosti raspodjela između populacija za ocjenjivana svojstva dlakavosti i oblika baze lista primijenjen je χ2 test homogenosti raspodjela.
Kako bi se istražila korelacija između fenotipskih svojstava provedena je standardizacija varijabli, te su Pearsonovi koeficijenti korelacije i statistička značajnost dobiveni pomoću funkcije 'cor.test' u R verzija 4.1.3 (R Core Team 2016).
Analiza glavnih komponenti (PCA) korištena je za kondenzaciju fenotipskih svojstava u nekoliko glavnih komponenti i istraživanje kontinuuma varijacije osobina. PCA je provedena korištenjem korelacijske matrice standardiziranih srednjih vrijednosti varijabli stabla. Analiza glavnih komponenti provedena je pomoću R paketa MorphTools2 u R verziji 4.1.3 (R Core team 2016) prema priručniku Koutecký (2015). Nadalje, konstruiran je dendrogram najbližih euklidskih udaljenosti na temelju metode neponderiranih parnih grupa s aritmetičkim sredinama (UPGMA) kako bi se provjerila struktura između proučavanih populacija. Euklidske udaljenosti određene su R funkcijom 'dist' korištenjem srednjih vrijednosti populacije standardiziranih metodom Z-score, a zatim su podvrgnute postupku klasteriranja (UPGMA metoda) korištenjem funkcije 'clust' u R paketu MorphTools2.
Analiza korelacije između mjerenih svojstava listova i geografskih i klimatskih faktora provedena je kroz izračun Pearsonovog koeficijenta korelacije.
REZULTATI - RESULTS
Fenotipska varijacija svojstava
Rezultati provedene statističke analize prikazani su u tablici 3. Za sva mjerena svojstva, koeficijent varijacije (CV) kretao se od 13.3% za duljinu plojke lista do 26.5% za duljinu peteljke lista.
Tablica 3. Zbirna statistika za 11 morfoloških svojstava proučavanih u 237 stabala Quercus petraea iz 24 prirodnih populacija u Bosni i Hercegovini
Table 3. Summary statistics of the 11 morphological characters studied in the 237 trees of Quercus petraea from the 24 natural populations in Bosnia and Herzegovina
CV – koeficijent varijacije (variation coefficient)
Fenotipska varijacija između i unutar populacija
Varijacije za 11 istraživanih fenotipskih svojstava mogu se podijeliti u 2 razine: između populacija i unutar populacija. Fenotipska svojstva listova bila su značajno (p < 0,001) različita unutar i među populacijama (Tablica 4). Srednja vrijednost koeficijenta diferencijacije populacija iznosila je 35,66%. Udio varijance između populacija kreće se od 8,3% za odnos duljine naspram širine lista do 30,8% za duljinu plojke lista. Udio varijance između stabala unutar populacija kreće se od 13,8% za udaljenost najšireg dijela plojke od baze plojke do 37,99% za duljinu peteljke lista.
Tablica 4. Udio komponenti varijance, koeficijenata diferencijacije populacije i F vrijednosti 11 svojstava na temelju svih podataka u 24 prirodne populacije Quercus petraea u Bosni i Hercegovini
Table 4. The proportion of variance components, population differentiation coefficients and F values of 11 traits based on all the data in 24 natural populations of Quercus petraea in Bosnia and Herzegovina
Rezultati testa homogenosti raspodjela između populacija prikazani su u tablici 5. Rezultati ukazuju na statistički visoko signifikantne razlike između populacija kada su u pitanju dlakavost lista i oblik baze lista (p<0.01).
Tablica 5. Rezultati testa homogenosti raspodjela između populacija za ocjenjivana svojstva
Table 5. Results of the test of homogeneity of distributions among populations for the assessed traits
Najveći broj listova nije imao dlakavosti (77,6%), po 1% listova imalo je dlakavost u pazuhu listova i na cijelom listu, a 21,4% listova je imalo dlakavost uz središnji nerv. Velik udio listova sa dlakavošću uz središnji nerv imale su populacije Bužim-Baštra, Ključ, Lašva-Desetnik, Srebrenik, Tjentište-Vrbnica, Zavidovići i Živinice-Litva.
Najveći broj listova imao je oblik baze lista 2 (73,3%), oblik baze lista označen sa 3 imalo je 22% listova, a oblik baze lista označen sa 1 imalo je 4,7% listova.
Korelacije između 11 svojstava lista
Pearsonov koeficijent korelacije korišten je za ispitivanje korelacija između 11 fenotipskih svojstava na srednjim vrijednostima stabla i prikazan na slici 5. Prvih 8 svojstava pokazalo je statistički značajnu pozitivnu korelaciju. Najveću pozitivnu korelaciju međusobno su pokazala svojstva NSDP i US (0,99) i NSLP i US (0,99), svojstva NSDP i NSLP (0,97), NSDP i UD (0,97), DPL i UNDP (0,86), DPT i OPP (0,82) i UNDP i UD (0.8). Najveću vrijednost negativnih korelacija međusobno su pokazala svojstva DPL i OSD (-0.36), UD i OSD (-0.34), UNDP i OSD (-0,31) i DPL i OPP (-0,25).
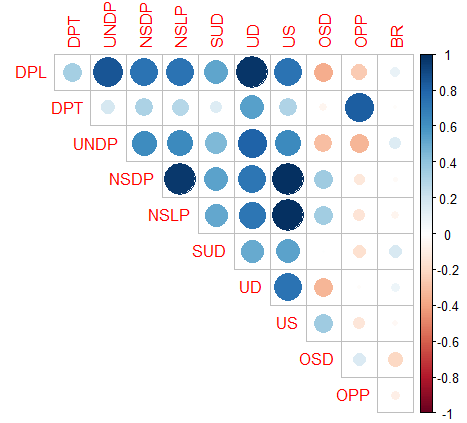
Slika 5. Analiza korelacije između 11 fenotipskih svojstava na razini stabla. Plava boja predstavlja pozitivnu, a narančasta negativnu korelaciju. Što je krug veći i boja tamnija, korelacija je jača (*p<0,05; **p<0,01; ***p<0,001). Fenotipska svojstava označena su skraćenicama prikazanim u tablici 3.
Figure 5. Correlation analysis among 11 phenotypic traits at the tree-level. Blue and orange colors represent positive and negative correlation, respectively. The bigger the circle and the darker the color, the stronger the correlation (*p<0.05; **p<0.01; ***p<0.001). Phenotypic traits are labelled by abbreviations shown in Table 3.
Glavne komponente i klaster analiza za 11 svojstava listova
Analiza glavnih komponenti korištena je za sumiranje varijacija među stablima u 11 morfoloških svojstava listova u neovisne smjerove varijacije (Tablica 6). Svojstvene vrijednosti prve tri glavne komponente bile su veće od 1, a zajedno su činile 88,8% varijacije. Glavni smjer varijacije (PC1, 58,2%) pokazao je negativne pondere za sva svojstva osim za odnos širine naspram duljine lista i odnos duljine peteljke naspram duljine plojke. Druga glavna komponenta (PC2, 17,9%) pokazala je pozitivne pondere za duljinu plojke lista, udaljenost najšireg dijela plojke od baze plojke, širinu usječenog dijela lista ispod najveće širine lijeve poluplojke i broj režnjeva sa desne strane. Treća glavna komponenta (PC3, 12,7%) pozitivne pondere pokazala je sa duljinom plojke, duljinom peteljke, širinom usječenog dijela lista ispod najveće širine lijeve poluplojke, ukupnom duljinom lista, odnosom duljine peteljke naspram duljine plojke i brojem režnjeva s desne strane.
Tablica 6. Vektori glavnih komponenti, svojstvene vrijednosti, stopa doprinosa i kumulativna stopa doprinosa prvih 11 glavnih komponenti na temelju srednjih vrijednosti stabla za 11 svojstava
Table 6. Principal component vectors, eigenvalues, contribution rates and cumulative contribution rates of first 5 principal components based on tree means for the 11 traits
Biplot konstruiran pomoću prve dvije glavne komponente prikazan je na slici 6. Na slici se uočava da se razlike između populacija vežu za skupinu morfoloških varijabli lista: UNDP, SUD, DPL, UD, NSLP i NSDP (PCA1) i skupinu OSD, OPP i DPT (PCA2). Najveće razlike u morfološkim svojstvima lista javljaju se između populacija Bihać-Gata i Fojnica sa značajno manjim dimenzijama lista (PCA1). U skupini populacija s većim dimenzijama lista, najveće razlike javljaju se između populacija Bihać-Gata i Kakanj-Begića Luka s najvećim DPT (PCA2).
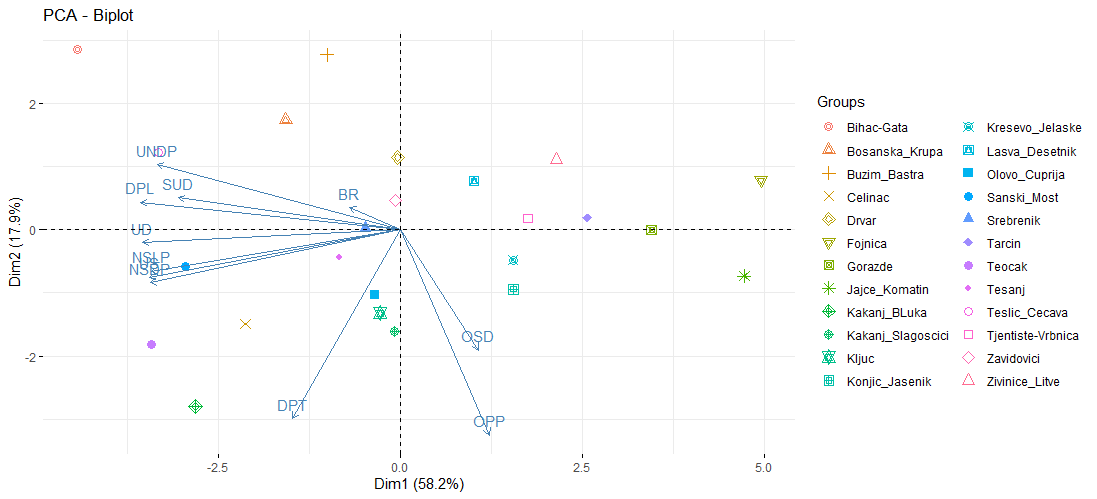
Slika 6. Biplot analize glavne komponente (PC) na temelju srednjih vrijednosti stabla za 11 morfometrijskih svojstava u 24 proučavane populacije Quercus petraea
Figure 6. Biplot of the principal component (PC) analysis based on tree means of 11 morphometric traits in the 24 studied Quercus petraea populations
Na temelju standardiziranih srednjih vrijednosti populacije za 11 morfoloških svojstava, 24 populacije se može podijeliti u dva klastera pomoću UPGMA metode (Slika 7). Prvi klaster se sastoji od populacija Fojnica i Jajce-Komotin, što se može objasniti manjom zemljopisnom udaljenošću između ovih populacija, a drugi klaster od svih ostalih populacija. Drugi klaster se sastoji od populacija koje su prilično nepovezane. Postoje poveznice klastera na nižim razinama koje se mogu objasniti, jer su populacije bliske, kao što su populacije Tarčin, Kreševo i Konjic, zatim populacije Sanski Most i Čelinac, te populacije Zavidovići i Tešanj. Klaster analiza pokazuje velik broj nelogičnih odstupanja, koja se vjerojatno mogu povezati s povijesnim događanjima u populacijama hrasta kitnjaka, kada je tijekom 19. stoljeća zajedno s hrastom lužnjakom intenzivno sječen (Begovic 1978, Memišević 2008). Tim sječama su značajno narušene populacijske strukture hrasta kitnjaka, te danas raspolažemo samo manjim dijelom nekada vrlo bogatog genofonda, jer su glacijalana pribježišta hrasta kitnjaka bila blizu područja koja istražujemo (Ballian i sur. 2007, Slade i sur. 2008).
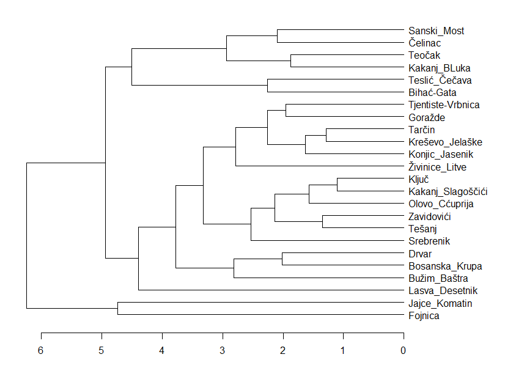
Slika 7. Odnos klastera 24 populacije Quercus petraea na temelju UPGMA metode i 11 morfoloških svojstava listova. Euklidska udaljenost korištena je za definiranje fenotipske udaljenosti između proučavanih populacija (morfološki podaci korištenjem populacijskih srednjih vrijednosti i standardizirani Z-score metodom). Brojevi uzorkovanih stabala svake populacije prikazani su u tablici 1.
Figure 7. Cluster relationship of the 24 Quercus petraea populations based on UPGMA method and the 11 leaf morphological traits. The Euclidean distance was used to define the phenotypic distance between the studied populations (morphological data using population means, standardized by the Z-score method). The sampled tree numbers of each population are shown in Table 1.
Korelacije između morfoloških svojstava listova i geografskih i klimatskih faktora
Rezultati korelacijske analize između mjerenih svojstava listova i geografskih i klimatskih faktora prikazani su u tablici 7. Vrijednosti Pearsonovog koeficijenta korelacije ukazuju na to da odnos duljine plojke i duljine peteljke lista značajno korelira s geografskim i većim brojem klimatskih faktora. Najveći broj analiziranih svojstava listova (osim duljine peteljke, odnosa širine i duljine lista, te broja režnjeva) u korelaciji je s geografskom širinom, te srednjom temperaturom kvartala s najviše padalina i količinom padalina u najtoplijem kvartalu.
Tablica 7. Pearsonov koeficijent korelacije između morfoloških svojstava listova i geografskih i klimatskih faktora
Table 7. Pearson's correlation coefficient between leaf morphological characteristics and geographical and climatic factors
DISKUSIJA - DISCUSSION
Fenotipske varijacije biljaka javljaju se kao rezultat djelovanja gena s jedne i faktora okoliša s druge strane, kroz dugi period evolucije (Miljković i sur. 2019) te se tako može reći da obilje fenotipskih varijacija u izvjesnoj mjeri može odražavati genetsku raznolikost istraživanog materijala (Wang i sur. 2023). Tako je fenotipizacija svojstava jedna od osnovnih metoda za istraživanje i ocjenu raznolikosti genetskih resursa šumskog drveća, a što je ključno za njihovo gospodarenje i očuvanje (Wang i sur. 2023). Nažalost, površina rasprostiranja bosanskohercegovačkih populacija hrasta kitnjaka je kroz povijest značajno reducirana (Begović 1978), kao i sama struktura stabala u populacijama, što se uz neadekvatan sustav gospodarenja odražava i na rezultate ovoga istraživanja.
Veličina i oblik lista također su rezultat interakcije više genetskih i okolišnih faktora (Gonzalez i dr. 2012), te fenotipske varijacije ovih svojstava nose informacije o genetskim i okolišnim varijacijama (Wright i sur. 2005, Niinemets i Sack 2006, Reich i sur. 2007, Niinemets 2015, Guet i sur. 2015). Veličina lista, morfologija, struktura i orijentacija izravno utječu na učinkovitost fotosinteze i imaju važan utjecaj na produktivnost (Marron i Ceulemans 2006), a oblik i veličina lista su adaptivna svojstva povezana sa svojstvima rasta i rani pokazatelj prinosa biomase (Marron i sur. 2007, Gebauer i sur. 2016). Također u istraživanjima koja su provedena na hrastu lužnjaku nađena je pozitivna korelacija između veličine lista i uzrasta kod potomstva (Ballian i Memišević Hodžić 2022) te time svojstva lista dobivaju na značaju.
U ovom istraživanju procjenjivali smo varijacije 13 fenotipskih svojstava listova hrasta kitnjaka iz 24 prirodne populacije u Bosni i Hercegovini. Utvrđena je fenotipska varijacija unutar i između populacija, ali je varijacija između stabala unutar populacije bila veća od varijacije između populacija za sva istraživana svojstva, kao što su za hrast lužnjak u Bosni i Hercegovini utvrdili Ballian i sur. (2010). Koeficijent varijacije svojstava kretao se od 11,9% za odnos širine naspram dužine lista (13% za ukupnu duljinu lista i 13.3% za duljinu plojke lista), do 26,5% za dužinu peteljke lista.
Provedena istraživanja na genetičkoj razini kod hrasta kitnjaka u Bosni i Hercegovini (Ballian i sur. 2007, Ballian i Bogunić, 2012, Ballian 2016), potvrđuju postojanje unutarpopulacijske i međupopulacijske varijabilnosti, kao što smo dobili i u ovom istraživanju. Nažalost ta istraživanja su obuhvatila samo manji broj populacija, te ne mogu ukazati na pravu sliku, za razliku od ovog istraživanja. U ovom istraživanju najveće vrijednosti koeficijenta diferencijacije utvrđen je za svojstva duljine plojke (49.16%), te širina usječenog dijela ispod najveće lijeve širine (48.46%). Prethodne studije hrasta kitnjaka koje su koristile molekularne markere pokazale su da se veći dio genetske varijacije nalazi unutar populacija, a manji dio između populacija (Gömöry i sur. 2001, Bruschi i sur. 2003, Ballian i sur. 2007, Ballian i Bogunić 2012, Ballian 2016, Rebrean i sur. 2023).
Također smo istraživali odnose između svojstava lista iz prirodnih populacija hrasta kitnjaka i geoklimatskih faktora te utjecaje tih faktora na fenotipsku varijaciju svojstava lista. Utvrđena je niska pozitivna korelacija, bez statističke značajnosti na razini povjerenja od 95%, između svojstava lista i analiziranih okolišnih faktora (geografskih i klimatskih varijabli zajedno). Arab i sur. (2021) također su utvrdili visok stupanj plastičnosti listova hrasta kitnjaka na atmosferske i pedosferske uvjete, te sugeriraju da hrast kitnjak ima visok potencijal kako bi se nosio s uvjetima rasta koji se očekuju kao posljedica budućih klimatskih promjena. Ovo tim više, jer ga u Bosni i Hercegovini nalazimo na sušnijim i skeletnijim staništima (Stefanović 1977, Stefanović i sur. 1983), a i dolazi na toplim tlima ofiolitske zone. Bruschi (2010) u rasadničkom testu šest populacija hrasta kitnjaka izloženih sušnom stresu utvrđuje znatnu diferencijaciju u srednjim svojstvima listova za sadnice proučavanih populacija, dok su zadržale nekoliko morfoloških razlika uočenih na terenu. Budući da su ove razlike u morfologiji pronađene u populacijama koje u rasadničkom testu rastu pod istim uvjetima, vjerojatno je da su registrirane razlike posljedica genetskih razlika. Međutim, nađen je visok stupanj fenotipske varijacije sa značajnim interakcijama populacija × način tretmana vodom. Općenito, Bruschi (2010) navodi da podrijetlo hrasta kitnjaka određuje varijacije u fenotipskoj plastičnosti kada se biljke uspoređuju pod različitim vodnim režimima. Naša dobivena varijabilnost pokazala je da je najveći broj svojstava u korelaciji sa srednjom temperaturom kvartala s najviše padalina te količinom padalina u najtoplijem kvartalu.. Tako u klaster analizi imamo povezivanje populacija koje imaju različite količine padalina, ali i razlike u njihovom godišnjem rasporedu. Ovdje se možemo i kritički osvrnuti na korištene podatke, koji nisu uvažavali mikrolokalne prilike u kojima raste hrast kitnjak. Stoga u budućim istraživanjima treba uvažavati i same mikroklimatska svojstva populacija. Primijenjena metoda se pokazala vrlo dobrom u istraživanju morfologije hrasta kitnjaka, te se može primijeniti i na druge vrste, uz dodatno uvažavanje mikroklimatskih svojstava populacija.
ZAKLJUČCI - CONCLUSIONS
Listovi hrasta kitnjaka iz 24 prirodne populacije iz Bosne i Hercegovine značajno su se razlikovali između i unutar populacija, a unutarpopulacijska varijabilnost bila je veća od međupopulacijske. Najviši koeficijent varijacije imalo je svojstvo duljine peteljke lista (26.5%), a najmanji duljina plojke (13.3%).
Najveći broj analiziranih svojstava listova (osim duljine peteljke, odnosa širine i duljine lista, te broja režnjeva) u korelaciji je s geografskom širinom te srednjom temperaturom kvartala s najviše padalina i količinom padalina u najtoplijem kvartalu.
Kako raspolažemo populacijama koje su se kroz povijest intenzivno sjekle, njihova genetska struktura je znatno izmijenjena, te u ovom istraživanju nismo dobili klinalnu varijabilnost koju smo očekivali, kada je riječ o nadmorskim visinama ili pravcu istok - zapad.
Dobivenu varijabilnost trebamo očuvati, jer će populacije hrasta kitnjaka kroz buduće generacije težiti uravnoteženom stanju, uz korekcije genetske strukture prirodnom selekcijom i migracijom.
Zahvala-Acknowledgments
Zahvaljujemo se Ministarstvu obrazovanja i nauke Federacije Bosne i Hercegovine za podršku projektu „Istraživanje unutarpopulacijske i međupopulacijske varijabilnosti hrasta kitnjaka (Quercus petraea (Matt.) Lieblein.) u Bosni i Hercegovini i izrada prijedloga mjera konzervacije“ kroz program financiranja naučno-istraživačkih projekata u Federaciji Bosne i Hercegovine za 2022. godinu, tokom kojeg smo prikupili potrebne podatke za pisanje ovoga rada. Zahvaljujemo se dr. Adnanu Hodžiću za učešće u aktivnostima projekta (istraživanje lokaliteta, herbariziranje, skladištenje i sušenje sakupljenog materijala te unos podataka), te studentima Nedimu Nogi (bachelor šumarstva), Selmi Huseinović (bachelor šumarstva) i Selmi Vehabović (bachelor hortikulture) za učešće u aktivnostima mjerenja prikupljenih uzoraka hrasta kitnjaka i unos podataka.
* ovaj odlomak nije lektoriran