
Commentary
Data on species plasticity and stable characters have an overall importance in identification keys: comments on Brullo et al. (2022) article
Gábor Sramkó 1,2, Attila Takács 1,2, Attila Molnár V. 1,2*, Agnieszka Popiela 3, Balázs András Lukács 4
1 University of Debrecen, Department of Botany, Debrecen, Hungary
2 ELKH-DE Conservation Biology Research Group, Debrecen, Hungary
3 University of Szczecin, Institute of Biology, Szczecin, Poland
4 Wetland Ecology Research Group, Centre for Ecological Research, Debrecen, Hungary
*Corresponding author e-mail: mva@science.unideb.hu
The genus Elatine L. consists of ephemeral species of wetland habitats that live in the temperate regions of both hemispheres. Because of their relatively fast life cycle and small habit, they lead inconspicuous lives, which is probably why they have been relatively little studied in European botany. Although the botanists of the 19 th and early 20 th century discovered all the currently recognised taxa, there were only a few studies that specifically addressed questions on their biology. This lack of detailed knowledge triggered the more recent interest in this genus (especially the European members), and several papers provided a huge amount of data and evidence about the species’ ecology, phenotypic plasticity, biogeography, karyology and molecular taxonomy.
In their recently published paper, Brullo et al. (2022) aimed to lectotypify Elatine macropoda Guss. and E. gussonei (Sommier) Brullo et al., two enigmatic members of the genus. They gave an overview of the taxonomic issue of these species, identified the type specimens, and gave a comprehensive description of both species, also aiming to clarify open questions in the nomenclature of these taxa. However, some of the points raised by Brullo et al. (2022) are in contrast to previously published scientific evidence, leading them to reach different taxonomic conclusions. In our view, this contrast is the result of (i) misinterpretation of some key findings published in our works, and (ii) adhering to preconceptions on the distribution and specific characteristics of these species. Given the importance of scientific discussion, we here attempt to shed light on contested points to help a better understanding of the taxonomy of this genus in Europe.
It was interesting to note that Brullo et al. (2022) reported hybridisation to be uncommon in Elatine, and suggested that it may be a rare phenomenon due to the prevalent autogamous nature of Elatine species. In their support of this statement, Brullo et al. (2022) cited Razifard et al. (2017), who reported the allopolyploid hybrid origin of E. americana (Pursh) Arn. and E. hexandra DC. in their work titled 'Reticulate evolution in Elatine L. (Elatinaceae), a predominantly autogamous genus of aquatic plants.' We acknowledge the reference made by Brullo et al. (2022) to Razifard et al. (2017) as an example of hybridisation in Elatine species. Furthermore, our own results demonstrated the presence of hybrid lineages in Elatine section Elatinella subsection Macropodae, which includes the focal species of the work of Brullo et al. (2022). Given our findings, along with the previous reports by Sramkó et al. (2016) and Takács et al. (2017), which were also cited by Brullo et al. (2022), it is possible that hybridisation in this genus may be more common than accepted. While the authors may have been aware of the presence of hybridisation in their focal group, we appreciate their analysis and interpretation of the available data, but we must disagree with them: hybridisation is not so rare in this genus.
In light of this, it is not appropriate to consider it a “surprising consequence” that two species of Elatine live in sympatry on the island of Sicily, because hybridisation – which does not seem as rare as suggested by Brullo et al. (2022) – requires the close encounter of different species at least at some point during their evolutionary history. It would have been more important in this respect to make a reference to the admixed lineage made up of E. gussonei from Lampedusa and Malta (Sramkó et al. 2016, Takács et al. 2017). The introgressed nature of these samples may explain some morphological differences of these populations from the rest of the distribution area of E. gussonei, which may be behind the view of seeing these populations as “different” by various scholars. In this respect it is noteworthy to refer to the unfortunate use of a Lampedusa plant without characterising its nuclear genome, as the lectotype of E. gussonei now selected by Brullo et al. (2022), can also be an introgressed plant.
Not only is there the problem of hybridisation, but our detailed study (Molnár et al. 2015) of the well-known phenotypic plasticity of vegetative characters in this amphibious genus (Mason 1956, Mifsud 2006) is another key piece in the literature that is overlooked by Brullo et al. (2022). Although they refer to the existence of phenotypic plasticity, in this context they failed to cite the results of Molnár et al. (2015) on the stability of seed characteristics and instability of vegetative and floral characters. This explains why they refer to the length of the petal and the sepal as one of the key characters separating the species E. macropoda and E. gussonei. Although Gussone (1827), Sommier (1907) and Pignatti (2017) emphasised the relevance of floral characters in the taxonomy of Elatine, we must take it into account that Mifsud (2006) has already documented the instability of these characters, which is simply rejected by Brullo et al. (2022) on the basis of a subjective evaluation (“ In our opinion, the floral traits cannot be linked exclusively to environmental conditions or flowers age”) and claim the opposite, citing their observations without measured and tested dataset (“ based on our observations, E. gussonei (Lampedusa and Malta) is morphologically distinct from the typical E. macropoda”). Sommier (1907) has already emphasized that E. gussonei differs from E. maropoda by its more curved seeds. In line with Sommier’s and Mifsud’s work, our measured dataset and statistical analyses demonstrate that seed morphology, especially its shape and surface ornamentation remains stable under different environmental conditions (Molnár et al. 2015), hence these are the most obvious morphological characters to differentiate species of Elatine, at least on the studied area. Moreover, this study also showed that the amount of light alone has a significant effect on the morphology of the vegetative and floral parts of the plants. Compared to plants growing under natural light conditions, the internodes, pedicels, caulin- and sepal leaves of in vitro grown individuals exposed to less intense artificial light are longer (Figs. 1. A, B).

Fig. 1. Flowering specimens of Elatine gussonei: A – cultivated ( in vitro) from Lampedusa, B – indigenous ( in situ) from Malta. Petal/sepal ratio is occasionally < 1. According to Brullo et al. (2022) erroneous assumption this is a distinguishing character that is specific to E. macropoda (see Brullo et al. 2022, Fig. 2). (Photo: B.A. Lukács).
Brullo et al. (2022) were selective in their choice when accepting the taxonomic importance of seed “ornamentation” (i.e., the shape of epidermal pits on the surface of the seed), but deny the utility of seed curvature, although our results (Molnár et al. 2015) clearly demonstrated the taxonomic value of this character (Fig. 2).
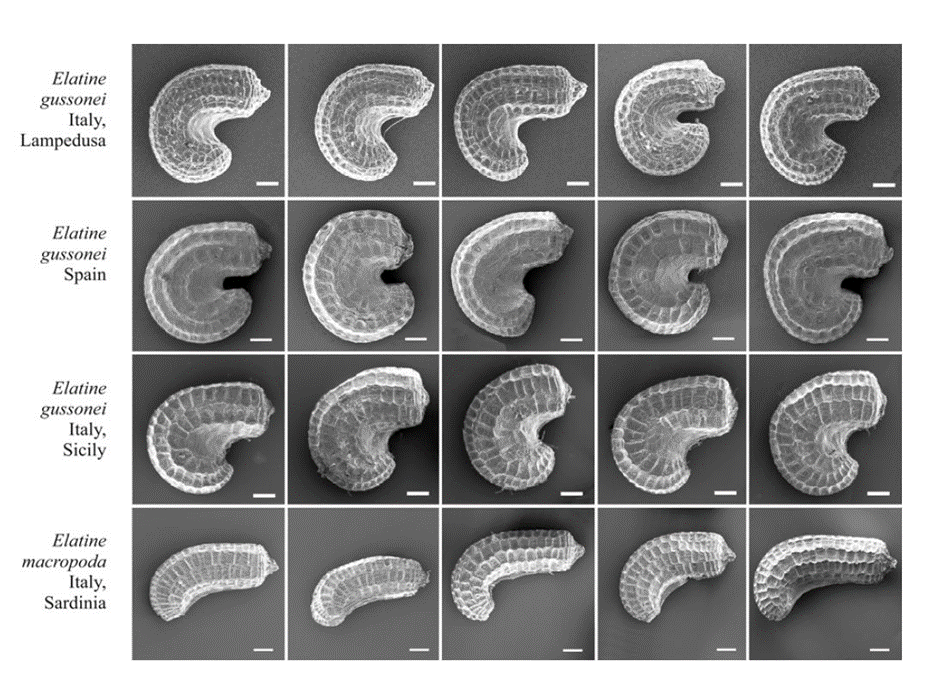
Fig. 2. Comparison of seeds of Elatine gussonei and E. macropoda. Scale bars = 0.1 mm. (SEM photo: A. Popiela).
It may be noted here that our very recently published paper (Łysko et al. 2022) emphasises this role even more: we tested several analytical methods on the discriminatory power of seed morphometry in the genus, where seed shape and ornamentation were found to be highly discriminatory. Regardless of this new result, however, Brullo et al. (2022) are wrong when they refer to Sramkó et al. (2016) as a source of information that seed morphology is a " quite variable trait even within the single populations" since this study summarised the seed morphology of different populations at the species-level (given the main goal of reconstructing the evolutionary history of the genus).
In order to demonstrate the usefulness of the preferred seed morphological characters, Brullo et al. (2022) published scanning electron microscope (SEM) images of seeds of E. macropoda (Brullo et al. 2022: Fig. 4) and E. gussonei (Brullo et al. 2022: Fig. 6), plus a comparative close-up image on epidermal pit shape of both species (Brullo et al. 2022: Fig. 5) where we can see pits of “rectangular or slightly hexagonal” shape as typical of E. macropoda (Brullo et al. 2022: Fig. 5A), and pits of “more or less isodiametric and usually hexagonal” shape as typical of E. gussonei (Brullo et al. 2022: Fig. 5B). Although the authors do not provide us with any measurement data for a statistically sound comparison, the visual inspection of their Fig. 4 would leave most observer with the impression that Figs. 4C and Fig. 4D (Sicilian plants from Modica and Ispica, respectively) are more similar to Fig. 5B, therefore, should be identified according to their epidermal seed pit shapes as E. gussonei.
In fact, both the seed curvature and epidermal structure clearly suggest the correct identification of the plants shown from Modica and Ispica as E. gussonei. Although Brullo et al. (2022) argue for the importance of longer petal length as a distinctive character that defines E. gussonei, it is rather easy to find Elatine plants with short petals on the island of Malta (see Fig. 1 B) where – according to Brullo et al. (2022) – only E. gussonei lives. Such plants were also presented by Mifsud (2006) and further examples can be seen in his photographic collection (https://maltawildplants.com/ELTN/Elatine_gussonei.php). If Brullo et al. (2022) were to consider the role of seed morphological characters in the genus and take into account the numerous publications that discuss the plastic nature of vegetative and floral characters (Molnár et al. 2014, 2015, Sramkó et al. 2016, Takács et al. 2017, Łysko et al. 2022) as well as the phylogenetic results (Sramkó et al. 2016, Razifard et al. 2017) in greater detail, they might find it easier to accept the presence of E. gussonei in Sicily and other Mediterranean areas. We must note here it is common in taxonomy for researchers to rely on different sets of characters and hold differing taxonomic opinions.
We accept, however, that this contradicts the well-established view on the very limited distribution and endemic nature of E. gussonei (Brullo et al. 1988, 2022), and would also necessitate the conservation re-evaluation of this species (Takács et al. 2017). Having said that we also think this species will still remain one of the key characteristic species of temporary Mediterranean ponds that – quite correctly! – enjoy the highest level of conservation interest in the European Union. Therefore, the taxonomic re-interpretation of E. gussonei and the consequently larger distribution area (it is still a Mediterranean endemic!) is not of concern for this plant from a conservation point of view. Instead, a better understanding of taxonomy is a fundamental prerequisite of well-established species conservation (Mace 2004).
In summary, Brullo et al. (2022) downplay i) the importance of seed shape as an identification character and ii) the environment- (primarily light intensity) and phenology- dependent nature of floral characters. While their results, which lack any report of detailed statistical analyses, are based on seed pit morphology, it is important to note that these values can be compared and verified. However, it is necessary to maintain scientific rigor, and the lack of detailed measurements and rigorous statistics in their report may limit the verifiability of the findings. Considering the contradictions between our previously presented coherent works and the recent claims of Brullo et al. (2022), we cannot accept their statements on the morphology and distribution of Elatine macropoda and E. gussonei. We further claim that their identification key for European (and not Mediterranean, as they indicated) Elatine species is misleading, since it focuses on phenotypically plastic characters and thus we recommend using the key presented in Popiela et al. (2017) to identify European species of Elatine.