
Introduction
The genus Placoneis Mereschkowsky was erected by Mereschkowsky in 1903 for a group of species showing a single chloroplast with a central bridge and lateral lobes (Mereschkowsky 1903). With the shift to an emphasis on using frustular features to diagnose taxa (e.g., Hustedt 1930), species assigned to Placoneis were considered part of the large genus Navicula Bory (1822). Due to its structure of the chloroplast, Cox (1987) resurrected the genus Placoneis with P. gastrum (Ehrenberg) Mereschkowsky (Basionym: Pinnularia gastrum Ehrenberg) as the type species. Phylogenetic analysis of Placoneis based on morphological and molecular data showed that the genus was part of the Cymbellales, a group with valves that are asymmetrical to the apical and/or transapical axes, despite having symmetrical valves. Support for this phylogenetic placement includes the straight and expanded central raphe endings and more or less hooked distal ones, striae composed of rounded areolae which are internally closed by volae (tectulum), and a single chloroplast with a central bridge and lateral lobes extending under the valves (Cox 1987, 2003, 2004, Mann and Stickle 1995, Bruder and Medlin 2007). To date, more than 136 species are recognized to be part of this genus (Guiry and Guiry 2022).
The genus Placoneis has a relatively wide distribution range, including Europe (Cox 1987, Bruder and Medlin 2007, Levkov and Williams 2011, Kulikovskiy et al. 2016, Vishnyakov 2020), Asia (Mayama and Kawashima 1998, Metzeltin et al. 2009, Kulikovskiy et al. 2012, Pomazkina et al. 2019, Kezlya et al. 2020), North America (Johansen et al. 2004, Kociolek and Thomas 2010, Kociolek et al. 2014), South America (Metzeltin and Lange-Bertalot 1998, Metzeltin and Lange-Bertalot 2007, Straube et al. 2013, Maidana et al. 2017), West Africa (Fofana et al. 2014) and Antarctica (Zidarova et al. 2009). In China, research on Placoneis has focused on the discovery of newly recorded species for the country, such as P. prespanensis Levkov, Krstic & Nakov (Li et al. 2010), P. explanata (Hustedt) S. Mamaya (Liu et al. 2012), P. interglacialis (Hustedt) E.J. Cox (Liu et al. 2012), P. opportuna (Hustedt) Chudaev & Gololobova (Lin et al. 2018), P. anglophila var. signata (Hustedt) Lange-Bertalot (Liu et al. 2020). New Placoneis species have been proposed from China by Gong et al. (2013) but otherwise only few new species of this genus have been reported. In Guangdong Province the diversity of diatoms is relatively rich (Qiu et al. 2016, Lin et al. 2018), but previous studies have mainly focused on Navicula Bory, Nitzschia Hassall, Cymbella C. Agardh and Gomphonema Ehrenberg (Wang et al. 2018, 2021). Our understanding of Placoneis from Guangdong Province is very limited (Wang et al. 2021).
The purposes of our study are: (i) to describe a Placoneis species new to science from the tributary of the Pearl River: Modaomen Channel, Zhuhai County, Guangdong Province, China based on detailed morphological observation using both light and scanning electron microscopy, (ii) to discuss its characteristics compared to related species, and (iii) to provide ecological information on this new species.
Materials and methods
Modaomen is located in Zhuhai County, Guangdong Province, China, one of the eight major gates at the mouth of the Pearl River. The length of the Modaomen Channel is about 45 km, and the water depth varies from 5 to 13 m. The Channel is relatively straight and about 2200 m wide (Chen et al. 2014, Tong et al. 2018).
In 2021, samples containing Placoneis were collected from the Modaomen Channel (22°24′20″N, 113°36′25″E). Channel water pH and specific conductance were measured using a YSI 650 multi-parameter display system (650 MDS, YSI Incorporated 1700/1725 Brannum Lane, Yellow Springs, OH 45387 USA) with a 600XL probe. The diatom samples were further processed as described by Battarbee (1986). After several rinses in distilled water, the partially cleaned diatom material was air-dried onto cover slips and mounted onto slides using Naphrax. The sample and slides were deposited in the Herbarium of the Institute for Ecological Research and Pollution Control of Plateau Lakes, Yunnan University, Kunming, P.R. China (YUK). The isotype slides were stored in the Key Laboratory of Biodiversity of Aquatic Organisms, Harbin Normal University.
Morphological observations of specimens were made under oil immersion at 1000× magnification with light microscopy (LM) using an OLYMPUS BX51-DIC research microscope and a C5060 Olympus digital camera. At least 500 valves were identified and counted in each surface sediment sample. Cleaned material for scanning electron microscope (SEM) analysis was air-dried onto cover glasses, mounted onto stubs, and coated with 20 nm of Au (EMSCOP SC 500 sputter coater). Resulting stubs were examined in the LEO 1530 scanning electron microscope (SEM). Description of the new species follows the terminology provided by Round et al. (1990), Cox (2003), Metzeltin et al. (2009) and Lange-Bertalot and Wojtal (2014).
Division Bacillariophyta Haeckel 1878: 95
Class Bacillariophyceae Haeckel 1878 emend D.G. Mann in Round et al. 1990: 651
Subclass Bacillariophycidae D.G. Mann in Round et al. 1990: 125
Order Cymbellales D.G Mann in Round et al. 1990: 653
Family Cymbellaceae Greville 1833: 263, 409
Genus Placoneis Mereschkowsky 1903: 3
Placoneis modaomenensis Y.-L. Li sp. nov. Fig. 1A–H; Fig. 1C is the holotype
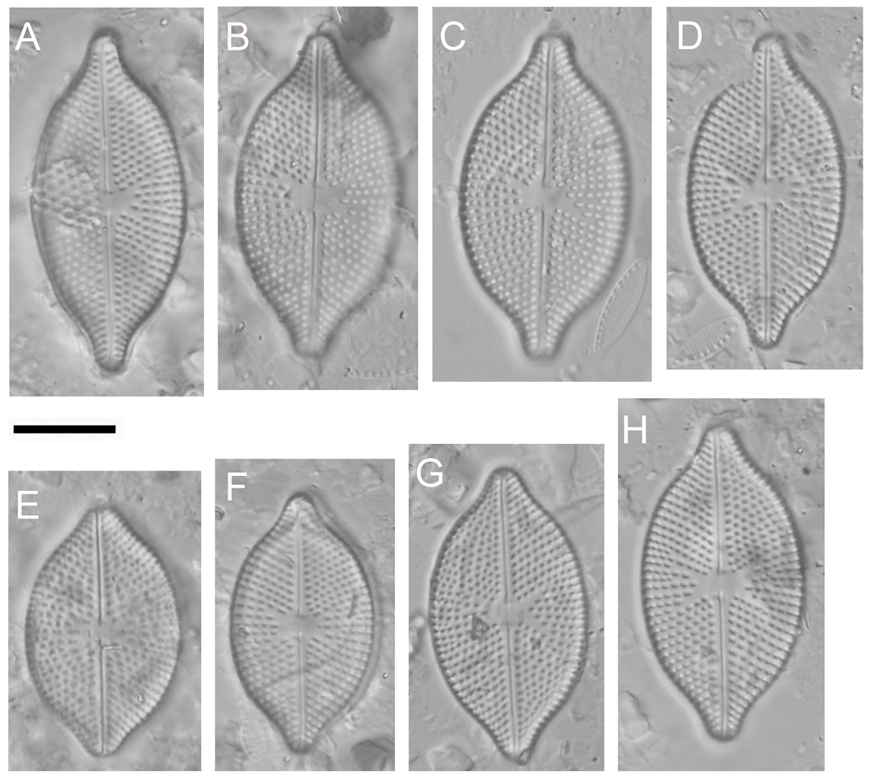
Fig. 1.Placoneis modaomensis, Light Microscopy (LM), Differential Interference Contrast (DIC). A–H – valve views, showing size range and variability of the holotype population. Scale bar = 10 μm.
LM (Fig. 1): Valves elliptical to broadly elliptical, nearly symmetrical about the apical axis, with rostrate or rostrate-rounded apices. Length 25.0–32.5 μm, width 14.0–16.5 μm, length/width ratio 1.62–2.16, median 1.89 (n = 30). Raphe filiform, almost straight with slightly expanded, but not clearly deflected to any side. External central raphe ends slightly straight, no significant expansion or bending. External terminal raphe fissures hooked to opposite sides. Axial area narrow, linear. Central area transverse, irregular to bow-tie-shaped, rarely asymmetrical, occupying nearly 1/2 of the valve width. Isolated pore is absent from the central area. Striae radiate throughout, 12–14 in 10 μm. Areolae visible, 11–14 in 10 μm.
In SEM (Fig. 2 and Fig. 3): External raphe narrow, central raphe endings hooked opposite to each other (Fig. 2A, B, G, H, I). The terminal fissures curved, deflected in the opposite directions, extend onto the valve margin (Fig. 2C, D, E, F). Striae uniseriate, composed of round or elliptical areolae, extending to valve margin (Fig. 2A–F). Areolae covered by volate occlusions (Fig. 2G, H, I). Each areola has almost 1–4 siliceous protrusions internally, forming a variety of different shapes (Fig. 2A–I).

Fig. 2.Placoneis modaomensis, Scanning Electron Microscope (SEM), external views. A, B – external view of an entire valve. C, D, E, F – valve apices, striae with elliptic/rounded areolae and hook-shaped terminal raphe fissures bent onto the valve margin. G, H, I – external view of valve center, the central raphe endings hooked in the opposite directions from each other, note volate occlusions. Scale bar = 2 μm.
Internal striae uniseriate, formed by elliptical areolae, separated by robust silica ribs (Fig. 3A–H). The raphe sternum clearly raised above the valve plane and expanded in the central area. Internal raphe straight, discontinuous with intermissio, lying in a prominent and raised raphe sternum. Central raphe endings straight, not expanded. Terminal raphe ends terminate as helictoglossae, offset from the raphe branch, bent slightly towards the valve margin (Fig. 3D, E, F). Striae are uniseriate, distinctly radiate, formed by round to elliptical shaped areolae (11–14 in 10 µm) and covered by dentate occlusions (Fig. 3G, H).

Fig. 3.Placoneis modaomensis, Scanning Electron Microscope (SEM), internal views. A, B, C – internal view of entire valve. D, E, F – internal view of valve apex with terminal raphe ends offset and bent slightly towards the margin. G, H – internal detail of central area showing raised central part and areolae covered by volate occlusions. Scale bar = 2 μm.
Type: – CHINA. Guangdong Province: Zhuhai County, Modaomen Channel, GD1, 22°24′20″ N, 113°36′25″ E, elevation 0 m a.s.l., samples collected by Dr. Hong-Qu Tang, 26 th July 2021. Holotype MDM202172601 in Coll. Yan-Ling Li, Yunnan University, Kunming, China. Fig. 1C is of the holotype; Isotype YUNGL20220218, Harbin Normal University, Harbin, China)
Etymology: – modaomensis, referring to the type locality from which the new species was obtained.
Associated diatom flora:Placoneis modaomensis is known from the Channel, situated at 0 m a.s.l. This species was associated with Amphora linearis F. Meister (1935: 97), Seminavis strigosa (Hustedt) Danielidis & Economou-Amilli (2003: 30), Aulacoseira granulata (Ehrenberg) Simonsen (1979: 58), Gomphonema parvulum (Kützing) Kützing (1849: 65), Navicula schroeteri F. Meister (1932: 38), Navicula viridula var. rostellata (Kützing) Cleve (1895: 15), Nitzschia clausii Hantzsch (1860: 40) and Nitzschia frustulum (Kützing) Grunow (1880: 98).
Ecology: The Modaomen Channel showed slightly alkaline conditions (pH 7.81), 29.8 °C of water temperature, 262 µS cm -1 of conductivity and 7.46 mg L -1 of dissolved oxygen (DO).
Discussion
Because Placoneis modaomenensis has symmetrical valve and radiate striae, it could easily be placed in one of four other genera: Paraplaconeis Kulikovskiy, Lange-Bertalot & Metzeltin (2012), Geissleria Lange-Bertalot & Metzeltin (1996), Rexlowea Kociolek & E.W.Thomas (2010) or Navicula Bory (1822). First, our new species resembles Paraplaconeis by symmetry of valve, but differs by the specific morphology of internal and external areolae patterns (Cox 1987, Cox 2003, Lange-Bertalot and Wojtal 2014). Second, P. modaomenensis is similar to the genus Geissleria on the basis of features observed with LM. These two genera are similar in terms of valve outline, but differ by the presence of the subpolar elongated areolae in the latter (Novais et al. 2013, Kulikovskiy et al. 2014). Third, P. modaomenensis is morphologically close to Rexlowea due to the valve outline, but these two genera are very distinct from one another by the arrangement, radiation and density of the striae (Kociolek and Thomas 2010). Fourth, P. modaomenensis can be confused with Navicula based on the valve symmetry. However, the former has exterior areolae covered by volate occlusions and internal areolae covered by dentate occlusions; while the latter areolae are all individually covered on the inside by a weakly convex hymen (Lange-Bertalot 2001, Li and Qi 2018). The characters found in Placoneis modaomensis are compared with those of morphologically most similar genera in Tab. 1.
While Placoneis seems easily placed as a member of Cymbellales, by virtue of its cytoplasmic features, its position in this lineage has been quite variable. Placoneis has been reported as outside the group of asymmetrical genera (Kociolek and Stoermer 1988, Nakov et al. 2014), deep within the cymbelloid lineage with some other genera naviculoid symmetry (Thomas et al. 2016, Kezlya et al. 2021) or in both positions depending upon the gene(s) used in the analysis (Bruder and Medlin 2007). Groupings within Placoneis are also enigmatic. Cox (1987) identified two groups within Placoneis, one with isolated pores, external terminal raphe fissures deflected in directions opposite one another and straight internal central raphe endings, while the other group has features of isolated pores absent, external terminal raphe fissures deflected in the same direction and recurved internal central raphe endings. Kociolek and Thomas (2010) noted some species from Colorado, USA, that did not conform to this organization of the genus. For example, P. fourtanierii Kociolek & Thomas (2010: 204) has external terminal raphe fissures that are deflected in opposite directions, but lack isolated pores and deflected internal raphe ends. P. coloradoensis Kociolek & Thomas (2010: 205) has external terminal raphe fissures that are deflected towards the same side, internal central raphe endings that are straight, but this species has isolated pores. In the case of P. modaomenensis, it has external terminal raphe fissures that curve in opposite sides, but has internal central raphe endings that are straight and no isolated pores. In addition, P. modaomenensis has areolae with volate occlusions positioned on the valve exterior which appears to be unique within Placoneis. Presence of central raphe endings turned opposite to one another in P. modaomenensis is shared with P. uruguayensis Metzeltin et al., a species without many features in common with other Placoneis species. The statement of Levkov et al. that “the full range of variation of morphology with the genus has been underestimated” (2007, p. 116) seems even more true now than when it was offered more than 15 years ago.
In terms of general valve shape and striae, P. modaomenensis shows some resemblance to P. amphibola (Cleve) E.J. Cox, P. amphiboliformis (Metzeltin, Lange-Bertalot and Soninkhishig) Vishnyakov, P. parvapolonica Lange-Bertalot & Wojtal, P. clementispronina Lange-Bertalot & Wojtal and P. nanoclementis Lange-Bertalot & Wojtal. However, P. modaomenensis differs strikingly from these species that are morphologically similar with respect to two features by the following: 1) external central raphe endings are bent in the opposite directions, and 2) both internal and external areolae are covered by volate occlusions.
Tab. 1. Morphological comparison of Placoneis to the most similar genera.
The characters found in P. modaomenensis are compared with other closely-related species of the genus in Tab. 2.
Based on valve outline, the new species is similar to P. amphibola, P. amphiboliformis, P. parvapolonica, P. clementispronina and P. nanoclementis, and P. modaomenensis sp. nov. most resembles P. parvapolonica based on its more convex valve margin and the structure of raphe and areola. However, they can be easily separated from one another. For example, in terms of the central raphe endings, the central raphe endings of the new species hooked in the opposite directions from each other while they do not clearly deflect to any side in P. parvapolonica. Comparing the shape of the areolae, the exterior areolae come in a variety of shapes and internal areolae are mostly round to elliptical in the new species, but P. parvapolonica differs in having areolae whose exterior and internal openings are nearly circular to slightly elongated in shape. Taking into consideration the occlusion of the areolae, the exterior and internal areolae are covered by volate and dentate occlusions respectively in the P. modaomenensis sp. nov., while there is an absence of occlusions in exterior and internal areolae in P. parvapolonica. Besides, the size range of P. modaomensis is larger than P. parvapolonica (16–20 × 8.0–9.5 μm). Finally, the stria and areola density are much lower compared to P. parvapolonica (stria: 14–15 μm, areola: 40–44 in 10 μm). In summary, these differences are sufficient to justify the description of P. modaomensis as an independent species.
Placoneis species are prevalent in freshwater bodies, including alkaline waters, mesotrophic and oligotrophic conditions (Cox 1987, Fujita and Ohtsuka 2005, Bruder and Medlin 2007, Kezlya et al. 2021). Of the five species similar to our new species, P. amphibola has a nordic-alpine distribution in Europe, and also occurs in freshwater fossil deposits (Cox 2003). Except for P. amphibola, the relevant information of nutrition, pH, and conductivity are not recorded, but other species have certain similarities or differences with our species. In terms of pH, P. parvapolonica, P. clementispronina and P. nanoclementis were found in alkaline waters, but P. amphiboliformis was known from acidic rivers (Metzeltin et al. 2009, Lange-Bertalot and Wojtal 2014). From the nutritional level, P. clementispronina and P. nanoclementis were distributed in eutrophic waters, and P. parvapolonica was found in mesotrophic waters (Metzeltin et al. 2009, Lange-Bertalot and Wojtal 2014). As for the conductivity, P. parvapolonica and P. nanoclementis are found predominantly in waters with moderate to high conductivity, whereas P. amphiboliformis was discovered rivers with low conductivity (Metzeltin et al. 2009, Lange-Bertalot and Wojtal 2014). Here, the new species of Placoneis was collected from alkaline waters with moderately high conductivity, which is the most similar to P. parvapolonica and P. nanoclementis.
In short, the discovery of this new species promotes the understanding of morphological features and ecological distribution about the genus Placoneis, and contributes to our understanding of diatom diversity, especially in Guangdong Province.
Tab. 2. Morphological comparison of the currently described species in Placoneis.