
Introduction
In Slovenia, Austria, Hungary and Croatia, pumpkin seeds ( Cucurbita pepo L.) are used to produce edible oil and its by-products can be used in ruminant feeding. The nutritional value of pumpkin seed is based on high energy potential, due to high percentage of oil (40 %-60 %; Bavec et al., 2007) and high crude protein content (30.8 %; Kim et al., 2012). Pumpkin seed cake is produced by pressing of hull-less seeds with screw press. This method of production preserves bioactive components, such as vitamins, provitamins, phytosterols, phospholipids and squalene, which are, together with some fatty acids, very important nutritional factors of pumpkin seed cake (Rabrenović et al., 2014). Due to the high content of digestible crude protein compared to soybean meal (496.9 vs. 396.0 g/kg) and high concentration of unsaturated fatty acids of which oleic (C18:1 n-9, OA) and linoleic (C18:2 n-6, LA) are predominant, pumpkin seed cake is a good quality feedstuff for goats (Zdunczyk et al., 1999). In livestock diets soybean meal was more frequently replaced by alternative sources of plant protein (Gugołek et al., 2017), due to its doubtful origin and quality (Hewlett and Azeez, 2008). Antunović et al. (2018) found that pumpkin seed cake can be used in lambs' feeding as a partial replacement for soybean meal in terms of blood parameters and carcass traits. Besides, the inclusion of pumpkin seed cake in ruminants´ diets has not been extensively researched yet.
On contrary, numerous studies reported a high nutritive value of extruded linseed added in concentrates mixtures for dairy goats regarding n-3 fatty acids in milk, especially α-linolenic acid (C18:3 n-3, ALA) (Klir et al., 2012; Nudda et al., 2013; Bernard et al., 2015; Klir et al., 2017). Production of milk that is low in medium-chain saturated fatty acids and high in odd and branched-chain fatty acids, C18:1 c9, and C18:2 c9t11 (CLA, conjugated linoleic fatty acid) and PUFA could have benefits on human health (Castro et al., 2019). Although, an excessive fat from linseed may have deleterious effects on digestive efficiency, milk fat and protein content (Doreau et al., 2015), while unsaturated fatty acids (UFA) in diets may lead to oxidative stress (Wang et al., 2010). One of the most important determinants of goats´ production efficiency is monitoring of herds' health status which is of fundamental importance for disease detections that impair goats' production (Rumosa-Gwaze et al., 2012). The beginning of lactation is the most demanding period in terms of energy balance status and also health management of herd (Walsh et al., 2011). High-yielding goats require high amounts of energy during early lactation leading animals to mobilize energy from their body reserves (Teh et al., 1994).
Therefore, the objective of the present study was to research if pumpkin seed cake in diets of early lactating dairy goats influences haematological and biochemical parameters in blood and milk quality as compared with diets containing soybean meal and diet where extruded linseed partially replaced soybean meal.
Materials and methods
Bioethics Committee for Research on Animals of Faculty of Agrobiotechnical Sciences Osijek established that the present research is carried out under the legal provisions according to Animal Protection Act of Croatia (NN 133/06, NN 37/13 and NN 125/13). Animal handling practices meets the European Union Directive 2010/63/UE about the protection of animals used for the experiment.
Experimental design, animals, and feeding
The research was conducted with 28 French Alpine dairy goats at family farm in Osijek-Baranya County (Croatia). All goats had kidded within a period of one week. After 7 days of adaptation period to experimental diets, animals were set to experiment (12 th ± 3 days of lactation). Duration of the current experiment was 20 days, during early lactation, which was a part of larger experiment lasting 75 days (Klir et al., 2017). The sampling was conducted when goats were approximately in the 32 nd ± 3 days of lactation. All goats were healthy and in adequate body condition (score 2.87 ± 0.51) according to Russel (1991) when the experiment started, and had an average age of 3.6 ± 1.73 years. Goats were machine milked twice per day, in the morning and the evening, and kept together in the barn. All goats were fed ad libitum with grass and red clover hay (60 % DM Phleum pratense and Lolium multiflorium, and 40 % DM Trifolium pratense). In addition, each goat was supplemented with 1 kg/day of a concentrate mixture individually. Goats were fed in separated feeding troughs during milking, and had free access to fresh drinking water, while no refusals of concentrate were observed. Dietary treatments differed in the main source of crude protein and crude fat in the concentrate mixtures. Control group contained soybean meal (n = 9), while in ELS group 38.3 % of soybean was replaced with ELS (n = 10), and in PSC group soybean meal was completely replaced with PSC (n = 9; Table 1). The diet had an approximate hay to concentrate ratio of 70:30 (on DM basis).
Table 1. Ingredient composition of the concentrate mixtures used in diets for early lactating goats
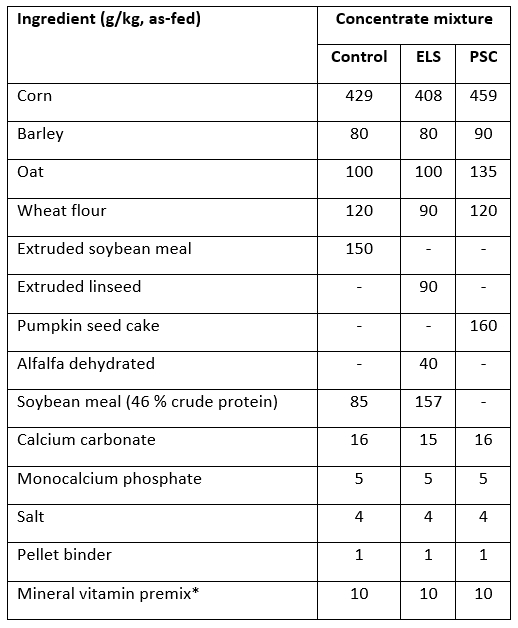
ELS - concentrate mixture with partial replacement of soybean meal with extruded linseed, PSC - concentrate mixture with complete replacement of soybean meal with pumpkin seed cake
*Mineral-vitamin premix: Fe (iron sulphate monohydrate) 4000 mg, Cu (copper sulphate pentahydrate) 800 mg, Mn (manganese oxide) 3500 mg, Zn (zinc sulphate monohydrate) 5000 mg, I (potassium iodide) 80 mg, Co (cobalt sulphate heptahydrate) 20 mg, Se (sodium selenite) 15 mg, Mg (magnesium oxide) 5000mg; vitamin A 1 000 000 IU, vitamin D3 150 000 IU, vitamin E (α-tocopherol) 1500 mg, vitamin K3 (menadione sodium bisulfite) 50 mg, vitamin B1 (thiamine mononitrate) 100 mg, vitamin B2 200 mg, vitamin B6 200 mg, vitamin B12 1 mg, niacin 1000 mg, Ca-pantothenate 500 mg, choline chloride 10 000 mg
Analysis of feedstuffs
Composition of feed was determined with standard methods (AOAC, 2006). The crude protein concentrations were estimated from nitrogen content according to the Kjeldahl method with the steam distillation unit for Kjeldahl nitrogen (Behr, Germany). The crude fat concentrations were analysed using the Universal Extractions System B-811 (Buchi, Switzerland) . Metabolizable energy (ME, MJ/kg DM) of feed samples were calculated from gas production after 24 h of in vitro incubation with Hohenheim gas test according to Menke and Steingass (1987). All plant samples were digested with 10 mL of a 5:1 mixture of HNO 3 and H 2O 2 at 180 °C for 60 min in microwave oven (CEM Mars 6, USA). The concentrations of mineral elements (Ca, P, Mg and Fe) in solutions of digested plant samples were determined by inductively coupled plasma mass spectrometer (ICP-MS, Agilent 7500a, Agilent Technologies Inc., California, USA). The value for N-free extract (NFE) was calculated as: NFE (g/kg DM) = 1000 - (crude protein + crude lipid + crude fibre + ash). Fatty acids in dietary ingredients were determined by gas chromatography following the methodology of State Office of Agricultural Chemistry in Baden Württemberg (LaChemie P23-5-008, V. 01).
Table 2. Chemical composition and major fatty acid proportions of hay and concentrate mixtures used in the diets for early lactating goats
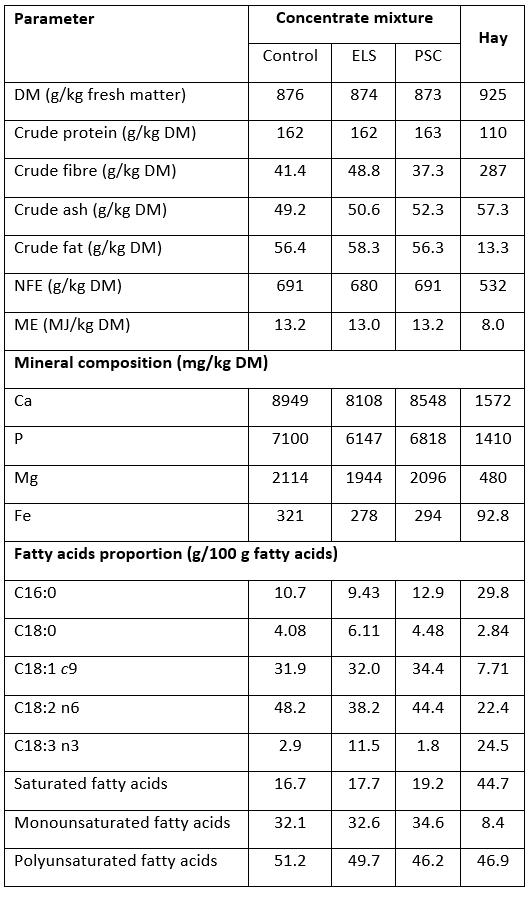
ELS - concentrate mixture with partial replacement of soybean meal with extruded linseed, PSC - concentrate mixture with complete replacement of soybean meal with pumpkin seed cake, DM - dry matter, NFE - nitrogen free extract, ME - metabolizable energy
Milk sampling and analysis
Two milk samples (2×100 mL) were taken on the 20 th day of experiment at 07.00 am from each goat by hand during morning milking. Immediately after sampling 100-mL milk sample from each goat was transferred into bottles containing 0.3 mL of the preservative azidiol, cooled to 4 °C and set up for analysis. Chemical composition was analysed by infrared spectroscopy HRN ISO 9622:2001 (Croatian Standards Institute, 2001) method on the analyser MilkoScan FT 6000 (Foss Electric, Denmark). Somatic cell counts (SCC) were determined by fluoro-opto-electronic method HRN ISO 13366-2/Ispr.1:2007 (Croatian Standards Institute, 2007) with a Fossomatic 5000 analyser (Foss Electric, Denmark), while the bacterial colony forming units (CFU) were determined by the method of epifluorescence flow ISO 16297:2013 [IDF 161:2013] (ISO, 2013).
Another 100-mL milk sample from each goat was frozen at -80 °C and stored until fatty acid analysis. The extraction of milk fat was done following a three-steps-extraction with gravimetric reference method (ISO, 2010). Fatty acid methyl esters were obtained from the milk fat samples by transesterification according to ISO (2002a). The fatty acid composition was determined by gas-liquid chromatography (ISO, 2002b). Fatty acids were calculated as weight percentage (g/100 g fatty acids) and are presented as means of duplicates.
Sampling of blood and analysis
The blood was collected from the jugular vein (10 mL) into sterile vacuum tubes Venoject ® (Sterile Terumo Europe, Leuven, Belgium) on day 20 after the morning feeding and milking. For haematology analysis, blood of goats was sampled into the sterile vacuum tubes containing ethylenediaminotetraacetic acid (EDTA) as the anticoagulant. Prior to analysis, whole blood was mixed with the Coulter mixer (Coulter electronics limited, Luton Bedfordshire, England). In whole blood, within 3 h after sampling, following haematological parameters were determined: number of leukocytes (WBC), erythrocytes (RBC), and the contents of haemoglobin (HGB), haematocrit (HCT), mean corpuscular volume (MCV), average haemoglobin content in erythrocytes (MCH) and mean haemoglobin concentration in erythrocytes (MCHC) on an automatic three differential haematology analyser (Sysmex PocH-100Iv, Sysmex Europe GmbH, Hamburg, Germany).
For biochemical analysis, within 4 h after sampling, serum (supernatant) in vacuum tubes was separated by centrifugation (10 min) at 1609.92 g and frozen at −20 °C until analysis. Within blood serum the concentrations of the minerals (calcium, phosphorus-inorganic, magnesium and iron), concentrations of urea, glucose, total proteins, albumin, cholesterol (CHOL), high density lipoprotein (HDL), low density lipoprotein (LDL), triglyceride (TGC), β-hydroxybutyrate (BHB) and non-esterified fatty acids (NEFA) as well as alanine aminotransferase (ALT), aspartate aminotransferase (AST) and gama-glutamyl-transferase (GGT) were determined. Globulin concentration was calculated as the difference between total protein and albumin. Analysis were obtained by Beckman Coulter (AU400, California, USA).
The activity of glutathione peroxidase (GPx) in serum was determined by Ransel® kits (Randox, UK) on an automatic analyser Beckman Coulter (AU400, California, USA) at a wavelength of 240 nm. The activity of total superoxide dismutase (SOD) in serum was determined by using RANSOD® kits (Randox, UK) on an automatic analyser Beckman Coulter (AU400, California, USA) at a wavelength of 510 nm.
Statistical analysis
A General Linear Model (GLM) was applied to the data for milk performance and biochemical parameters, with diet treatment as fixed effect. Means were compared using the Tukey test and differences between the groups were declared significant at P < 0.05. Values of SCC and CFU were logarithmically converted to a linear score with the aim to approximate normal distribution. Correlations between parameters of goats´ energy status (BHB and NEFA) and selected milk fatty acids and ratios were evaluated by Pearsons´ correlation with CORR procedure. Statistical analysis was performed using the statistical software SAS (2013, SAS Inst. Inc., Cary, NC. version 9.4).
Results and discussion
Milk yield and chemical composition of goat milk were not affected by the feeding treatment (Table 3). Similarly, the soybean replacement by PSC or partially by ELS did not result in any significant differences regarding the SCC and CFU. This indicated that PSC can be used as an alternative to soybean for the conditions of this study. Both PSC and ELS have a different fatty acids profile compared with soybean, but those changes in the diet did not affect either milk production. However, extruded linseeds addition in goats' diets lowered the milk yield, and increased the milk fat content in the study conducted by Chilliard et al. (2013).
The milk fatty acid profile has been changed when feeding goats with ELS and there were minor changes in PSC compared to Control (Table 4). The ELS and PSC diets did not influence the proportions of C6:0, C8:0 and C10:0, fatty acids responsible for the particular sensory properties of goats´ milk (Bessa et al., 2016) and health benefits providing better digestibility of goat milk (Zenebe et al., 2014). Bernard et al. (2015) found that SCFA, specifically C6:0, C8:0 and C10:0, were lowered in milk when feeding goats with 216 g/kg DM of extruded linseed in goats´ diets compared to control diets. The LCFA inhibit lipogenic enzymes of the mammary gland, leading to lowering of de novo synthesized fatty acid proportions (Chilliard et al., 2007). In the present study the proportion of ELS in the concentrate mixtures was probably not high enough to influence a lower proportion of these SCFA. Contrary, proportion of branched-chain fatty acids (BCFA) like C14:0 iso, and C15:0 iso, were significantly higher (P<0.05) when feeding goats with ELS compared to Control, while in PSC remained intermediate and unaffected. Supplementation of ruminants´ diet with different lipid source has relevant effect on ruminal bacteria community in goats (Cremonesi et al., 2018). Abuelfatah et al. (2016) reported that inclusion of linseed in the diet of goats at level of 10 or 20 g/100 g DM increases the molar proportion of acetate in rumen, which may be the reason for the increase of above stated milk fatty acids. Besides, Goetsch (2019) explained that numerous factors are responsible for microbial population in the rumen of dairy goats, including greater feed intake relative to body weight and shorter time of digesta residence in the rumen, and some other effects before treatment.
Table 3. Milk yield and chemical composition and fatty acid proportions in milk of early lactating goats fed with diets containing extruded linseed (ELS) and pumpkin seed cake (PSC)
SEM - standard error of mean; SCC - somatic cells count, CFU - number of bacterial formed colonies; a,bRow means with different superscripts differ significantly at P<0.05
Table 4. Fatty acid proportions in milk of early lactating goats fed with diets containing extruded linseed (ELS) and pumpkin seed cake (PSC)
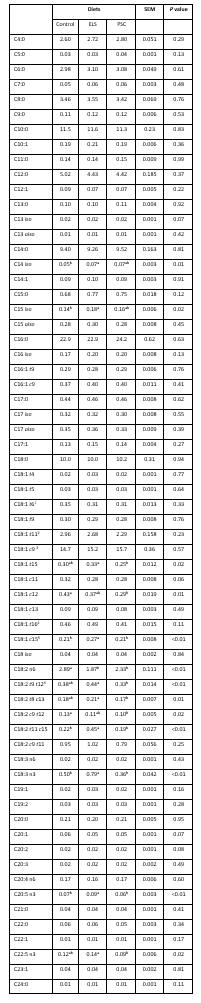
1Contains C18:1 t7 + t8 as a minor components, 2Contains C18:1 t10 as a minor component, 3Contains C18:1 t12 + t13 + t14 as a minor components, 4Contains C18:1 c14 as a minor component, 5Contains C19:0 a minor component, 6Contains C18:2 t9 t12 + t8 c12 + c9 t13 a minor components; SEM-standard error of mean; a,bRow means with different superscripts differ significantly at P < 0.05
Proportions of stearic (C18:0), OA, vaccenic (VA, C18:1 t11) and CLA remained unaffected by dietary treatments. Bernard et al. (2015) determined that addition of extruded linseed in combination with fish oil may be efficient in increasing VA and CLA proportions in milk. Although proportions of CLA, which is of the interest with respect to human health, were appropriate as compared to another studies, the amount of CLA consumed to improve human health still needs to be established (Hur et al., 2017).
In the present study ELS was abundant in ALA, which resulted in higher proportions of biohydrogenation metabolites, like C18:1 t15, C18:1 c15 and C18:2 t11 c15 compared to Control and PSC. Shingfield et al. (2010) reported that within putative pathway of ALA biohydrogenation C18:1 t15 and C18:1 c15 are produced as minor fatty acid metabolites, while in major ALA biohydrogenation pathway C18:2 t11 c15 is formed. Similar was reported in milk of ELS compared to Control and PSC of the present study.
Diets with PSC and ELS lowered LA compared to the Control, as a consequence of the higher LA concentration in the Control feed mixture containing soybean meal and extruded soybean (Table 1). This was reflected in lowered total n-6 fatty acids in ELS and PSC (Table 5). Proportion of ALA was the highest in ELS which resulted in higher ALA and total n-3 proportions, and lower LA/ALA ratio in milk of ELS goats compared to those of Control and PSC. The relationship between ALA intake and ALA proportion in milk is linear and transferred from diet to milk is close to 4.5 % (Doreau et al., 2015). According to the Commission Regulation EU (2010), a claim that food is a source of omega-3 fatty acids, may only be made when the product contains at least 0.3 g of ALA per 100 g, while food is high in omega-3 fatty acids when the product contains at least 0.6 g ALA per 100 g. As a consequence of greater ALA supply from the ELS diet proportion of EPA was higher compared to Control or PSC, as ALA is a precursor of EPA during the elongation.
Table 5. Groups of fatty acids in milk from early lactating goats fed with diets containing extruded linseed (ELS) and pumpkin seed cake (PSC)
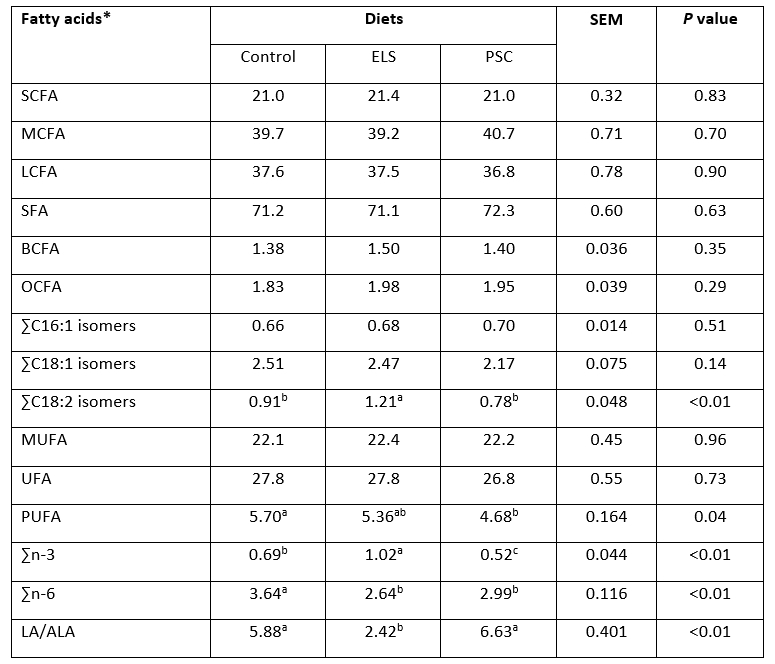
*SCFA - short-chain fatty acids, MCFA - medium-chain fatty acids, LCFA - long-chain fatty acids, SFA - saturated fatty acids, BCFA - branched-chain fatty acids, OCFA - fatty acids with odd number of carbon atoms, MUFA - monounsaturated fatty acids, UFA - unsaturated fatty acids, PUFA - polyunsaturated fatty acids, LA - linoleic acid, ALA-α - linolenic acid; SEM - standard error of mean; a,bRow means with different superscripts differ significantly at P < 0.05
The concentration of globulins was the highest when PSC was fed and the lowest for the Control, while the rest of haemato-chemical parameters in goats’ blood resulted unchanged by the diets (Table 6). This observation could be due to high phenolic compounds in pumpkin seed cake of PSC diet as reported by Peričin et al. (2009). Polyphenol rich fraction from food have been shown to result in a significant increase in the IgM- and IgG-plaque forming cell responses of blood leukocytes (Hikosaka et al., 2007). The addition of feedstuff rich in crude fat in ruminants´ feeding may affect the production traits and health of ruminants (Nudda et al., 2013). All values of haemato-chemical parameters were within values determined in lactating Alpine goats by Antunović et al. (2013 and 2017).
Table 6. Haemato-chemical parameters from early lactating goats fed with diets containing extruded linseed (ELS) and pumpkin seed cake (PSC)
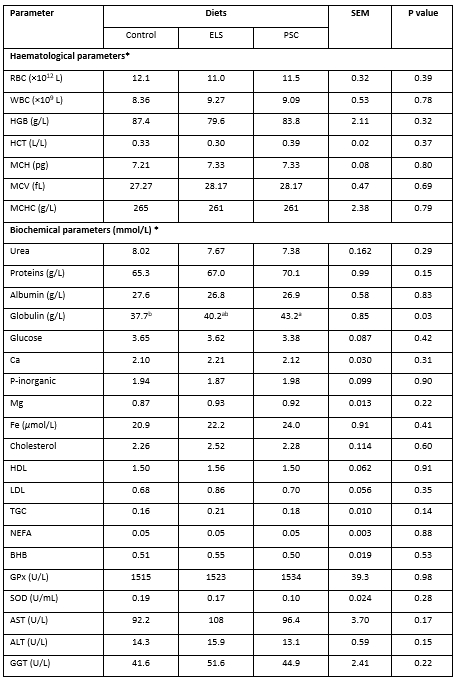
*RBC - erythrocytes, WBC - number of leukocytes, HGB - contents of haemoglobin, HCT - haematocrit, MCH - average haemoglobin content in erythrocytes, MCV - mean corpuscular volume, MCHC - mean haemoglobin concentration in erythrocytes, HDL - high-density lipoprotein, LDL - low-density lipoprotein, TGC - triglycerides, NEFA - non-esterified fatty acids, BHB - β-hydroxybutyrate, GPx - glutathione peroxidase, SOD - superoxide dismutase, AST - aspartate aminotransferase, ALT - alanine aminotransferase, GGT - gamma-glutamyl transferase, SEM - standard error of mean; a,bRow means with different superscripts differ significantly at P<0.05
It is well known that ruminants are susceptible to oxidative stress during pregnancy and lactation (Bernabucci et al., 2005), resulting in physiological state of metabolic stress that may cause diseases by changing the metabolic status (Antunović et al., 2013). The concentration of BHB in blood (≥0.8 mmol/L) and changes of the body score index (≤2.0) are specific indicators for assessment and prevention of metabolic disorders of highly productive lactating goats (Binev et al., 2014). Jahani-Moghadam et al. (2015) did not determine significant differences in the glucose, NEFA, BHB and urea concentrations in blood of cows fed ELS. Likewise, the results of the current study indicate the positive energy balance in early lactating dairy goats fed with ELS or PSC.
The ELS or PSC did not influence the activity of enzymes of antioxidant status. Bernabucci et al. (2005) reported that oxidative status is associated with energy status, which means that higher concentrations of NEFA and BHB are associated with lower activity of antioxidative enzymes in plasma. Additionally, Wang et al. (2010) established lower activity of SOD in plasma of cows fed UFA rich diets compared to cows fed with diets added SFA, while GPx activity and BHB concentration did not differ between groups. Owing to similar concentration of UFA in diets (Table 2) and stable energy status (Table 6), differences in the activity of the SOD and GPx enzyme were missing, and the antioxidant status was preserved.
Negative correlation was determined between the NEFA and the total SCFA or certain SCFA (Table 7). Short chain fatty acids which are synthesized de novo in the mammary gland decrease when plasma NEFA concentration increases (Craninx et al., 2008). The variability of milk C18:0 and OA concentration is linked to changes in the energy balance during the first 4 months of goats' lactation, since ruminant adipose tissue is very rich in palmitic, stearic, and OA (Chilliard et al., 2003). Established correlations result from the negative effect of LCFA mobilized from adipose tissue on de novo synthesized SCFA (Barber et al., 1997), as confirmed by present study on goats. Möller et al. (2018) determined the relationship between MCFA in hair and fat metabolism of cows. Dórea et al. (2017) found high sensitivities of milk C6:0, C8:0, C10:0 and C12:0 related to NEFA concentrations in cows' plasma. Accordingly, these authors determined threshold values to detect high NEFA concentrations in plasma, but when the proportion of these fatty acids are above these thresholds, the cow is classified as low concentrations of NEFA. In the present research a negative correlation was determined between the serum NEFA and C10:0 as well as C15:0 in goat milk while the other individual SCFA did not show any significant differences. Dórea et al. (2017) established that plasma NEFA concentrations may be predicted by linear regression models using rations C18:1 to C15:0 and C17:0 to C15:0 in cows´ milk. In the present study Pearson correlation showed positive correlation between both ratios in goat milk and serum NEFA, and only C17:0 to C15:0 ratio and BHB in serum. These correlations indicate low mobilization of lipids from adipose tissue in early lactating dairy goats in the current research.
Table 7. Correlations between the selected milk fatty acids and parameters of energy status in the serum of early lactating goats
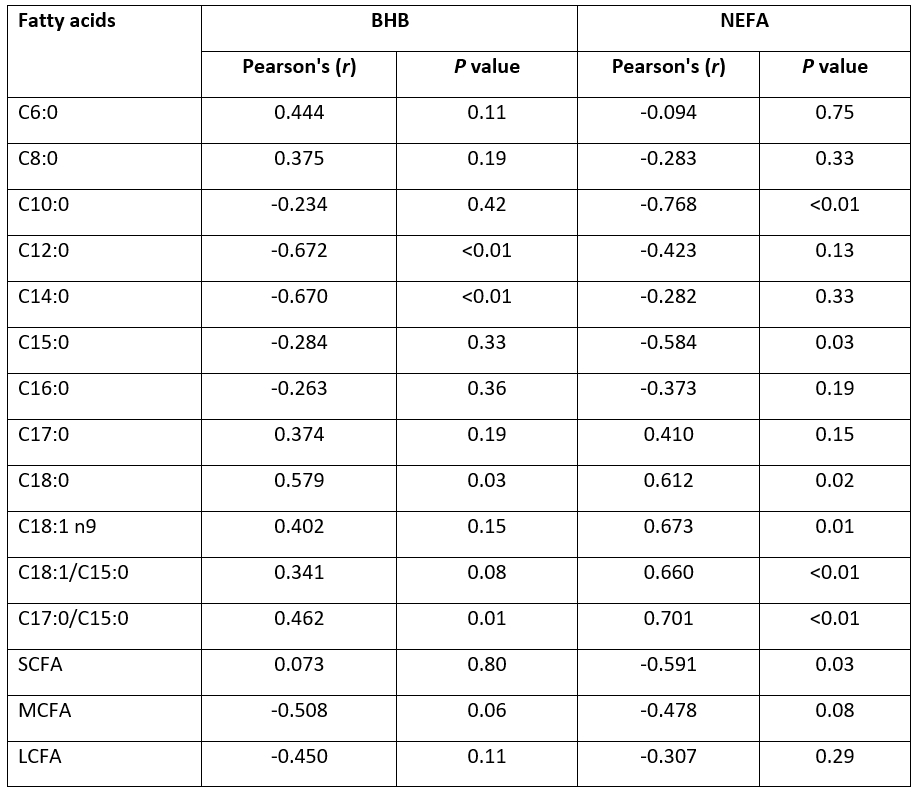
NEFA - non-esterified fatty acids, BHB-beta-hydroxybutyrate, SCFA - short-chain fatty acids, MCFA - medium-chain fatty acids, LCFA - long-chain fatty acids
Conclusion
The current findings indicate that the replacement of soybean meal with pumpkin seed cake (PSC) or partially with extruded linseed (ELS) is feasible with respect to haemato-chemical parameters of the early lactating dairy goats. Moreover, these indicate adequate health, preserved antioxidant and nutritional status when goats were fed with PSC or ELS. Partially substituting soybean meal with ELS enhanced goat milk with ALA and total n-3 fatty acid proportions while compared to conventional diets, soybean meal replaced by PSC lowered linoleic acid in milk. In further study it is necessary to determine antioxidant status of milk which will contribute to better understanding whether PSC is suitable for goats´ feeding regarding milk quality.
Acknowledgements
The study was carried out within the research team Innovative breeding and technological processes in animal production (No. 1126) at Faculty of Agrobiotechnical Sciences Osijek.
Utjecaj pogače sjemenki bundeve i ekstrudiranog lana u obrocima na hematološko-biokemijske pokazatelje u krvi i na kvalitetu mlijeka alpske koze tijekom rane laktacije
Sažetak
Zbog visokog sadržaja sirovih bjelančevina i visoke koncentracije nezasićenih masnih kiselina, pogača sjemenki bundeve (PSC) potencijalno je kvalitetno krmivo za preživače, dok je ekstrudirani lan (ELS) u obrocima koza već pokazao blagotvoran utjecaj na kvalitetu kozjeg mlijeka. Laktacija je vrlo zahtjevno razdoblje za praćenje metaboličkog statusa životinja. Prema tome, cilj ovoga istraživanja bio je utvrditi utjecaj PSC i ELS na hematološko-biokemijske pokazatelje u krvi i na kvalitetu mlijeka, kao zamjena za soju u obrocima mliječnih koza. Istraživanje je provedeno s 28 koza pasmine francuska alpine koje su bile podijeljene u dvije pokusne skupine tijekom 20 dana u ranoj laktaciji. Obroci su bili izbalansirani u pogledu energije i sadržaja sirovih bjelančevina, uz neznatne razlike sadržaja lipida. Obroci kontrolne skupine (n = 9) sadržavali su sačmu i ekstrudiranu soju. U obrocima pokusnih skupina soja je djelomično zamijenjena s ELS (n = 10) ili u potpunosti zamijenjena s PSC (n = 9). Hematološko-biokemijski pokazatelji u krvi koza nisu rezultirali značajnijim promjenama pod utjecajem hranidbe, iako je obrok s PSC povećao koncentraciju globulina u serumu. PSC je rezultirao smanjenjem linolne kiseline (n-6) u odnosu na kontrolnu skupinu. ELS obroci povećali su koncentraciju α-linolenske, eikozapentaenske kiseline i koncentraciju ukupnih n-3 masnih kiselina u mlijeku. Utvrđene su pozitivne korelacije između ne-esterificiranih masnih kiselina (NEFA) i C18:1/C15:0, NEFA te C17:0/C15:0, kao i β-hidroksibutirata te C17:0/C15:0. Potpuna zamjena sačme i ekstrudirane soje s PSC ili djelomično s ELS opravdana je s obzirom na hematološko-biokemijske pokazatelje koza u ranoj laktaciji, dok je ELS imao blagotvoran utjecaj na masne kiseline u mlijeku u odnosu na PSC.
Ključne riječi: pogača sjemenki bundeve; ekstrudirani lan; mliječne koze; hemato-biokemijski pokazatelji; kvaliteta mlijeka
References
https://doi.org/10.1016/j.aninu.2016.10.004
https://doi.org/10.5194/aab-60-409-2017
http://dx.doi.org/10.4314/sajas.v48i4.11
https://doi.org/10.24099/vet.arhiv.0780
https://doi.org/10.3168/jds.S0022-0302(05)72878-2
https://doi.org/10.1017/S1751731114003048
https://doi.org/10.1590/1678-457X.05315
https://doi.org/10.3390/ani9050205
https://doi.org/10.3168/jds.S0022-0302(03)73761-8
https://doi.org/10.1016/j.anifeedsci.2013.08.002
https://doi.org/10.3168/jds.2007-0656
https://doi.org/10.1017/S1751731117003433
Croatian Standards Institute (2007): HRN ISO 13366-2/Ispr.1:2007 (ISO 13366-2:2006)
Milk-Enumeration of somatic cells-Part 2: Guidance on the operation of fluoro-opto-electronic counters, Croatian Standards Institute, Zagreb, Croatia
Dórea, J.R.R., French, E.A.,Armentano, L.E. (2017): Use of milk fatty acids to estimate plasma nonesterified fatty acid concentrations as an indicator of animal energy balance. Journal of Dairy Science 100, 6164-6176.
https://doi.org/10.3168/jds.2016-12466
https://doi.org/10.1051/ocl/2015042
https://doi.org/10.1080/09712119.2019.1580585
https://doi.org/10.1080/1745039X.2017.1322796
https://doi.org/10.1002/ptr.2033
https://doi.org/10.1016/j.livsci.2016.11.016
https://doi.org/10.3168/jds.2014-8789
https://doi.org/10.4162/nrp.2012.6.1.21
https://doi.org/10.1017/S175173111700060X
Menke, K.H., Steingass, H. (1987): Schätzung des energetischen Futterwerts aus der in vitro mit Pansensaft bestimmten Gasbildung und der chemischen Analyse. II. Regressionsgleichungen. Übers Tierernährung 15, 59-94.
https://doi.org/10.1017/S0022029918000791
https://doi.org/10.1017/S1751731113000931
https://doi.org/10.1016/j.foodchem.2008.07.079
https://doi.org/10.1016/j.lwt.2013.10.019
https://doi.org/10.3168/jds.S0022-0302(94)76948-4