
Introduction
Changes in plant community composition following both primary and secondary succession have been studied on a global scale. Some global-scale ecological patterns during successional changes are common and can be detected in many different biomes (see Prach and Walker 2020). However, purely floristic changes during succession can be highly dependent on the region under study, as they depend on the local species pool, land-use history and disturbance type, as well as on environmental filtering effects. Therefore, floristic changes can differ even within a single study area of the same climate and same climax community. This can make it hard to formulate generalizations about the floristic changes during succession, except in the cases when generalizations are made for a specific area of interest. In Croatia, vegetation succession is poorly studied. Only rare data are available on grassland succession (Krstonošić et al. 2016, Kutnjak 2010, Randić 2007), whereas somewhat more comprehensive research on the whole successional sequences has been carried out in the neighboring country of Slovenia (Čarni et al. 2007, Čarni et al. 2021, Dakskobler 2010). In this paper, we depict the local-scale community composition changes following secondary succession of oak-hornbeam forest after cessation of agricultural land-use (i.e. grassland abandonment) in NW Croatia.
Study area
The study was conducted in NW Croatia, in the surroundings of the village Brlog Ozaljski (45° 37' 32.37" N, 15° 24' 11.09" E). The sessile oak and common hornbeam forest (association Epimedio- Carpinetum betuli) is the dominant vegetation cover of the area. The soil type is calcocambisol on limestone. In the study area, during the last few decades, the abandonment of agricultural land use led to a progression of vegetation succession, resulting in a significant loss of meadow and pasture communities. Today, the remaining meadows are used as hay pastures, i.e., for obtaining hay and for low-intensity rotational grazing by sheep.
Vegetation survey
The sequence of vegetation succession was studied using the space for time substitution, and five stages of succession were included in the study (Fig. 1).
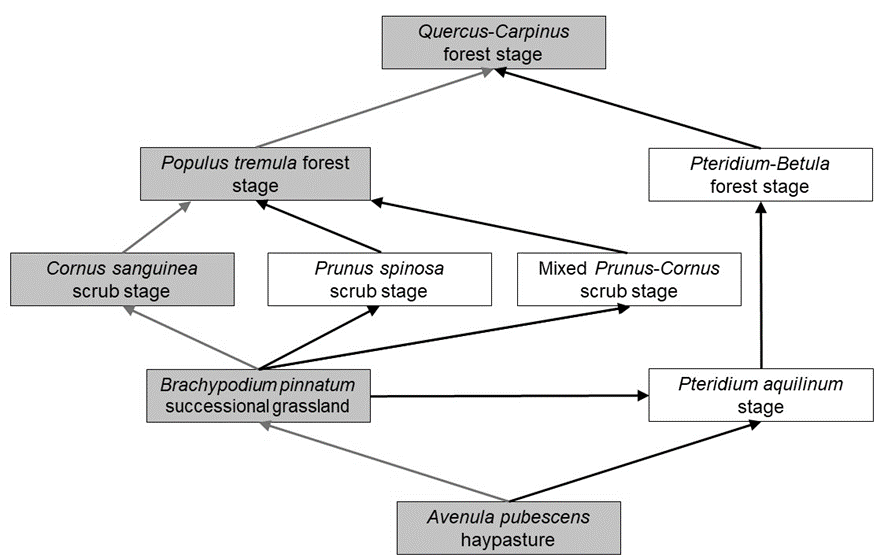
Fig. 1. Different successional pathways of oak-hornbeam forest development after cessation of agricultural land-use (i.e. grassland abandonment) observed in the study area. The successional sequence marked in grey was analysed in the present study.
The sequence was determined during preliminary field investigations. In the last century, the lands of the study area that are today hay pastures were used in various ways, mostly as extensive arable fields. The age of succession since abandonment was roughly estimated with preliminary field investigations, imagery from Google Earth Pro 7.3.6. time series and Croatian Geoportal (https://geoportal.dgu.hr/), and by interviews with a local farmer. The approximately estimated age of the studied successional stages following hay pasture abandonment was as follows: successional grasslands (2–5 years), shrub stages (5–15 years), Populus tremula L. stage (15–30 years), and oak-hornbeam stage (> 30 years). The vegetation survey was performed throughout 2019 and 2020 in May, June, July, and August, using five 100 m 2 plots (10 × 10 m) within each successional stage, totaling 25 plots. Species cover in grasslands was recorded using the extended Braun-Blanquet scale, and in scrub and forest stage using the classical Braun-Blanquet scale. Nomenclature of plant taxa followed POWO (https://powo.science.kew.org/).
Data analysis
Differences in community composition between the successional stages were plotted with non-metric multidimensional scaling (NMDS) using a species × site matrix based on Bray-Curtis dissimilarities. This was done in PAST 4.03 software. Prior to the NMDS, cover values of species occurring in more than one vegetation layer were pooled, and all of the cover values were transformed to type I ordinal percentage scale (Van der Maarel 2007).
Results and discussion
The NMDS ordination plot (Fig. 2) provided a very good representation of the community composition differences between the successional stages.
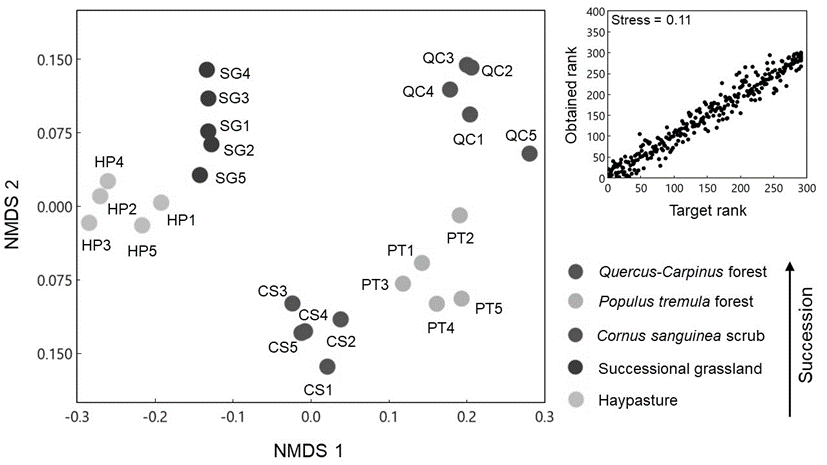
Fig. 2. Non-metric multidimensional scaling ordination (NMDS) of successional stages based on community composition differences obtained by Bray-Curtis dissimilarities. The plot in the upper right corner represents the Shepard plot. The direction of succession progression is well represented by the first two NMDS axes.
The succession direction was in correspondence with the first NMDS axis. Total number of species found in the present study was 160, whereas total species number observed in hay pasture, successional grassland, C. sanguinea scrub, P. tremula forest, and oak-hornbeam forest was 76, 72, 59, 46, and 59, respectively. The plant community composition of successional stages is listed in On-line Suppl. Tab. 1.
1. Spread of Brachypodium pinnatum (L.) P. Beauv. after hay pasture abandonment
After cessation of hay pasture management, Brachypodium pinnatum completely colonized the habitat, reaching a cover of > 75%. These successional grasslands seem to be a part of the order Brachypodietalia pinnati Korneck 1974. The only frequent species in B. pinnatum grassland were Dactylis glomerata L., Briza media L., Filipendula vulgaris Moench., and seedlings of Cornus sanguinea L. and Prunus spinosa L. Even though the number of species after the spread of B. pinnatum remained somewhat high, it decreased the abundance of many hay pasture species, some of them completely ceasing after it had spread. Due to its incredible colonization efficiency attributed to clonal propagation, but also efficient generative dispersal (Bąba et al. 2012), B. pinnatum spread is considered a threat to plant biodiversity of native calcareous grasslands in Europe (Bobbink and Willems 1987). It can decrease species diversity through light reduction by its dense canopy and litter accumulation (Bobbink and Willems 1987), and its dense root system is likely to be efficient in competing for nutrients. On the other hand, its spread is a natural part of the succession after cessation of agricultural land use in many areas of Croatia. However, it provides low-quality forage due to high silica levels in its tissues and high amount of structural C (Canals et al. 2017), thus, its spread in our study area has a negative impact on agronomic quality of local hay pastures.
2. Cornus sanguinea scrubs (CS)
C. sanguinea gained a complete dominance in this stage, with a cover of 90–100%. Almost all species in the herb layer persisted with a cover of <1%. As its fruits are dispersed by frugivorous birds in high densities, it can easily dominate after cessation of land management (see Kollmann and Grubb 2001). The most frequent species under C. sanguinea bushes were Erigeron annuus (L.) Desf, Centaurea jacea L. , Ajuga reptans L. , Clinopodium vulgare L. , Fragaria vesca L. , Veronica chamaedrys L., Prunus spinosa, and Rubus fruticosus L. This stage would likely correspond to the ass. Corno-Ligustretum Horvat ex Trinajstić & Z. Pavletić 1991. Rubus fruticosus had a somewhat high overall abundance as it was able to intertwine its stems around the branches of C. sanguinea, thus acquiring more sunlight. Carpinus betulus L. seedlings already appeared in the herb layer of this stage, but B. pinnatum and Populus tremula were not present in our plots. We noticed that the shrub stages were full of wildlife routes, mainly that of rabbits, roe deer, and wild boars, which might be responsible for the suppression of the herb layer. For P. tremula, another possibility is that the saplings were not yet present in the shrub stage, and their growth may occur sometime later by cloning via suckers of the nearby trees (Worrell 1995).
3. Late-successional Populus tremula forest stage (PT)
P. tremula was dominant in the tree layer, accompanied by Betula pendula and Carpinus betulus. Cornus sanguinea remained present in the shrub layer, but its dominance was low compared to that in the scrub stage. Corylus avellana L. was also abundant in some plots. In the herb layer, C. betulus seedlings were frequent, and those of Cornus sanguinea and Prunus spinosa still survived in these shaded conditions. Epimedium alpinum L. and Hedera helix L., the species specific for climax forest, were already present in this stage. It could be considered that this stage resembles some form of ass. Populo tremulae- Betuletum pendulae (Glišić 1950) Trinajstić 2004, even though the uniqueness of this association is questionable (Vukelić 2012), as it probably highly depends on the successional context.
4. Late-successional oak-hornbeam forest stage (QC)
Carpinus betulus was the dominant tree in this stage, accompanied by Quercus robur L . In the shrub layer, only Corylus avellana appeared in four plots, whereas in the herb layer, Epimedium alpinum, Rubus hirtus Roxb., and the seedlings of C. betulus occurred in all plots. R. hirtus occasionally occurred in the previous successional stages, but became frequent in the herb layer of this stage, whereas E. alpinum, a species of Illyrian-Balkan chorotype diagnostic for the Illyrian oak-hornbeam forests, dominated the herb layer. This stage was only an approximation to the climax of Illyrian oak-hornbeam forests (ass. Epimedio-Carpinetum betuli) that were once present in the study area.
5. General remarks on the studied succession
Species survival throughout succession differed. Ajuga reptans and Fragaria vesca survived in all stages. These two species are known to have a high ecophysiological plasticity to changes in environmental conditions, especially in relation to light availability. A. reptans can grow well in high light exposure, but in shaded conditions such as under the forest canopy, it can maintain itself likely due to efficient use of nitrogen from the humus in order to increase chlorophyll synthesis, and consequentially, light capture (Golovko and Dymova 1999). Under canopies, F. vesca is known to allocate more energy to aboveground biomass production at the expense of root biomass (Hancock and Bringhurst 1978), which can lead to increase of leaf area in order to expand light capture efficiency. Viola hirta was not present in hay pastures, but it did appear in successional grasslands and persisted throughout the succession. Veronica chamaedrys and Clinopodium vulgare survived all the way to the Cornus sanguinea scrub stage. In the herb layer, C. sanguinea and Prunus spinosa, being highly dispersed by frugivorous birds, persisted from successional grassland to the late-successional Populus tremula stage.
Conclusion
In the studied area, after hay pasture management is abandoned, succession begins with the spread of Brachypodium pinnatum, after which many hay pasture species decrease in abundance or completely disappear. Succession further proceeds to a Cornus sanguinea-dominated scrub stage, followed by a forest stage with Populus tremula as a dominant tree. Succession ends with an oak-hornbeam forest (association Epimedio-Carpinetum betuli). Only five generalist species survived in almost all succession stages.