
Introduction
For a long time, fungi have been overlooked in the invasion science literature, apart from the fungal pathogens that have strong negative impacts on biodiversity and the economy (e.g. in the agriculture). Symbiotic fungi have been to some extent surveyed, but saprotrophic fungi rarely (Desprez-Loustau et al. 2007, 2010). According to Desprez-Loustau et al. (2007), lack of the quantification of the impacts of non-pathogenic alien fungi may be a major reason for the scarcity of surveys and the data on them. In last 15 years, a number of European countries have compiled lists of alien fungi (see Voglmayr et al. 2023, for countries and references), but as shown from the example of Austria in which they expanded their first checklist (Voglmayr and Krisai-Greilhuber 2002) with an almost five-fold increase in the second edition (Voglmayr et al. 2023), it is an ongoing process. As highlighted by Voglmayr et al. (2023) plant pathogens are the dominant ecofunctional group in the list of Austrian alien fungi, which is the case in other countries as well. Saprobionts are still generally underrepresented in alien fungi lists. It should be noted that biogeography of fungi is much less known and clear than that of plants and animals (Desprez-Loustau et al. 2007).
Among the alien saprotrophic fungi in Europe, Clathrus archeri (Berk.) Dring is one of the better known, with lots of papers reporting on its distribution across Europe (see the review by Parent and Thoen 1986, and Parent et al. 2000). One reason for this may be the peculiar appearance of its sporocarps (Fig. 1), making it easier for identification, and attracting more attention, even from non-fungi experts and the general public.

Fig. 1.Clathrus archeri observed by authors in November 2023 in Donji Stupnik (left image – eggs; middle image – sporocarp) and Donje Ladanje (right image – sporocarp). Pen in the left image is 14 cm long. Photo: Sven D. Jelaska.
According to Parent and Thoen (1986) the first record in Europe of this fungus, originating from Australia and New Zealand, is from 1920 in Vosges, France (Lemasson 1923 in Parent and Thoen 1986), where it is believed to have most probably arrived with wool products. From there it gradually spread all over Europe. It has been recorded in great variety of habitats including different types of forests (deciduous, coniferous – natural and plantations, mixed coniferous-deciduous), grasslands, orchards, disturbed habitats, gardens (Parent and Thoen 1986, Bîrsan et al. 2014, Pietras et al. 2016, Brännhage and Gross 2020). However, although it can be found in a diversity of habitats it does prefer humid and acidic soil (Parent and Thoen 1986, Pietras et al. 2016).
Published data on C. archeri in Croatia are extremely scarce and to our knowledge, with respect to the scientific literature, these are: Tkalčec et al. (2005) in which the species is mentioned for Croatia for the first time in the scientific literature; Kranjčev (2008) who reports on its presence in two locations in northwest Croatia; Pietras et al. (2021) who included in their analysis one piece of data (out of four) on the fungus’s presence in Croatia deposited on GBIF.org on October 26th, 2018. Apart from the scientific literature, C. archeri was included in all three editions of the “Ordinance on Protection of Fungi” (Official Gazette 1998-2002), but was omitted from the subsequent “Ordinance on Designation of Wild Taxa as Protected or Strictly Protected” (Official Gazette 2006) and “Red Book of Croatian Fungi” (Tkalčec et al. 2008). Preliminary inclusion of C. archeri on the list of protected fungi was probably a result of a lack of data and information on the fungus’ origin, which happened in other countries as well, in the Eastern part of Europe where C. archeri arrived and was observed later (e.g. Romania – Tănase and Pop 2005, Bulgaria – Gyosheva et al. 2006, Ukraine – Didukh 2009). Increased knowledge and data collected resulted in its removal from the Red Book of Ukraine (Heluta et al. 2022).
To fill the current gap in the knowledge of the presence and distribution of C. archeri in Croatia, we collected reliable available data on its presence, based on which we analysed the ecological preferences of the species. Furthermore, we developed habitat suitability models to get some indication on its potential overall distribution in Croatia, which can be also used for planning future research on the species.
Material and methods
To gather information on the presence of C. archeri in Croatia we have searched for fungus’s records in GBIF.org (https://www.gbif.org/); iNaturalist.org (https://www.inaturalist.org/), Mushroomobserver.org (https://mushroomobserver.org/) and Wood (2008). Furthermore, we have used several query terms in different combinations (“Clathrus”, “archeri”, “Anthurus”, “Devil’s fingers”, “Octopus stinkhorn”, “velika polipovka”, “Croatia”, “Hrvatska”) in the following sources:
- Portal of Croatian scientific and professional journals (https://hrcak.srce.hr/en)
- Croatian Research Information System (https://www.croris.hr/)
- Scopus (https://www.scopus.com)
- Internet web pages
Queryings scientific literature sources revealed just two publications in which C. archeri was mentioned: Tkalčec et al. (2005) and Kranjčev (2008). Only the latter gave locations (two) for the fungus in Croatia. GBIF.org did not contain all the localities present in iNaturalist, because of licensing restrictions assigned to some observation records. We have contacted owners of these observations and obtained permission to use their observations in this analysis. They are presented in Tab.1 with corresponding links, while data present in GBIF on April, 24th 2024 can be found in the GBIF.org (2024) downloaded data set.
Tab. 1. Localities of Clathrus archeri in Croatia with WGS84 coordinates, month and year of observation, and source of data (all web sources accessed on April 24, 2024.).
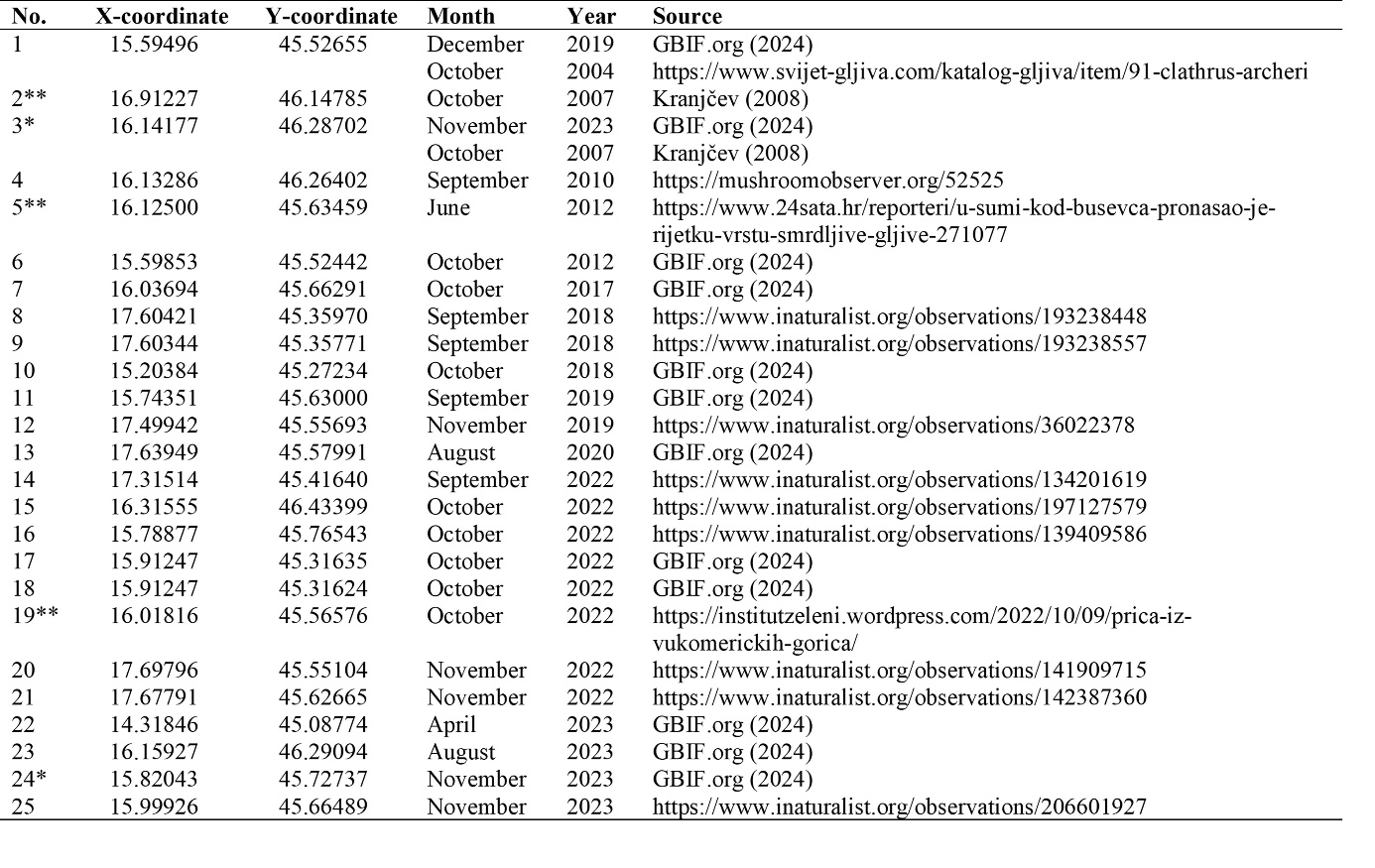
From the Internet web pages search, we have included three results (No. 1, 5, and 19 in Tab. 1) that mentioned month and year of observation, in addition to a locality that could be georeferenced. For such location in the Karlovac City area (No. 1 in Tab. 1) there was an observation in GBIF.org (2024) with exact coordinates, hence we used that location but kept the web source presented in Tab. 1 for being among the first data of the presence of the fungus in Croatia. Descriptions of two locations of C. archeri in Kranjčev (2008) were sufficiently precise in just once case, the presence of which we confirmed in November 2023 (No. 3 in Tab. 1) and assigned coordinates using the Global Navigation Satellite System (GNSS). Overall, we have synthesized data on 25 locations of C. archeri in Croatia (Fig. 2) with the oldest data originating in 2004.
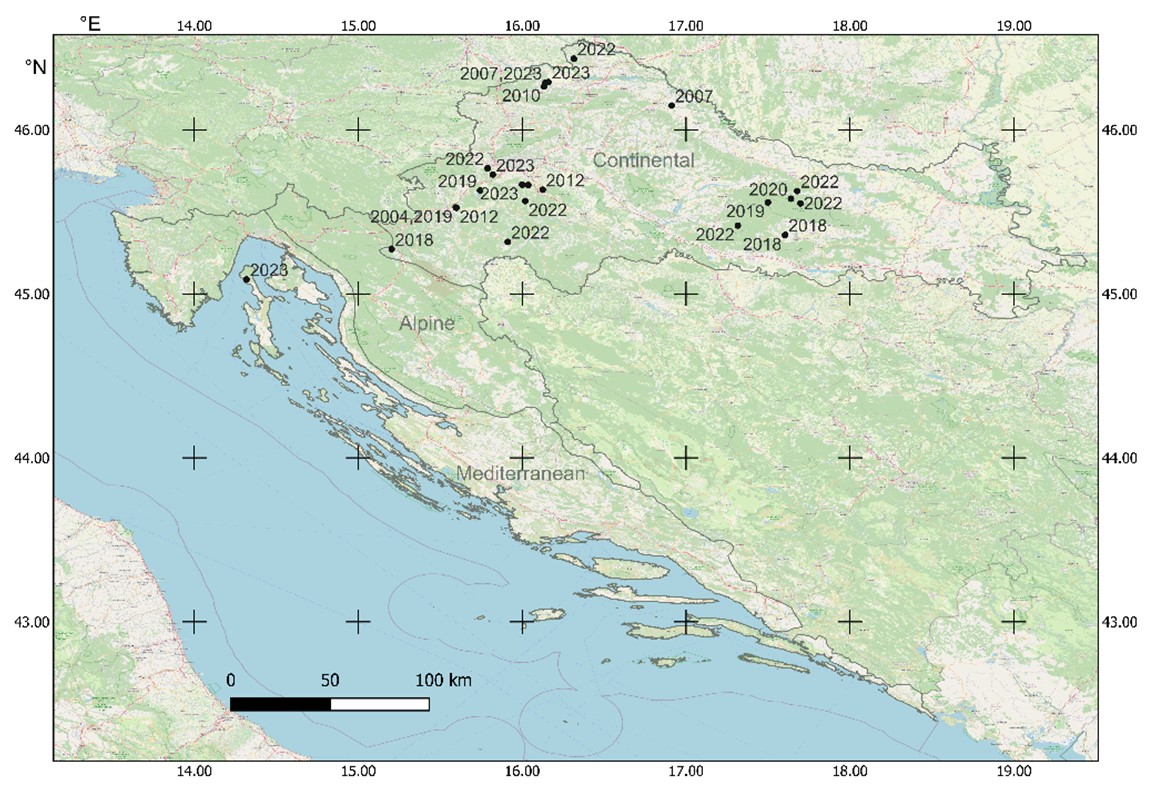
Fig. 2. Distribution of Clathrus archeri in Croatia (details of localities in Tab. 1), with NATURA2000 biogeographical regions indicated. Background map is OpenStreetMap (Base map and data from OpenStreetMap and OpenStreetMap Foundation (CC-BY-SA). https://www.openstreetmap.org and contributors)
To develop habitat suitability models, we used Maxent software (Phillips et al. 2024), 22 data with exact coordinates on the occurrences of C. archeri in Croatia (Tab. 1) and two sets of environmental variables. In the first iteration, we used eight climatic variables (mean temperature and amount of precipitation per meteorological season) based on a 30-year period (1960-1990, Croatian Meteorological and Hydrological Service). In the second iteration, in addition to climatic variables, pH value (in H2O) of the topsoil (0-5 cm depth) was used. For latter we used a dataset available for Europe (Poggio et al. 2021). All environmental variables were prepared as 300 m resolution grids. Given the number of occurrence data, we have used all as a training set. Consequently, only Area Under Curve (AUC) for the training set was available. Three localities to which exact coordinates could not be assigned (No. 2, 5, and 19 in Tab. 1), were used as an independent test data set to evaluate the habitat suitability models developed.
In addition to climatic and pH variables mentioned above, we assigned values of elevation and slope (based on a 300 meter resolution digital elevation model of Croatia) to all 25 localities, and calculated several descriptive statistical measures (minimum, maximum, mean, 10th and 90th percentile) to get some insight into the ecological preferences of C. archeri in Croatia. Elevation and slope were not included in habitat suitability modelling since elevation is correlated to climate (positively with precipitation, and negatively with temperature) and value of slope can vary significantly on short distances, and hence could increase uncertainty of the habitat suitability model.
At two locations (No. 3 and 24 in Tab. 1, Donje Ladanje and Donji Stupnik respectively) where we observed C. archeri in November 2023 we collected soil samples. After removing the leaf litter, upper 5 cm of the soil was sampled with a cylindrical corer (8 cm in diameter) in triplicates around the observed sporocarp (with a distance of 1 m between the subsamples) and stored as a composite sample representative for the location. Samples were transported to the laboratory where they were air dried at room temperature. To measure pH values, a combined electrode pH meter (HANNA HI 99121, Direct soil pH meter) was used on suspensions composed of 10 g of soil and 25 mL of distilled water. Prior to measuring, the suspensions were covered for 30 min with occasional stirring. Organic matter content was determined by annealing in a muffle furnace (INKO LP-08). According to the NRM Laboratories methodology (Davies 1974) ground soil samples were first dried in an oven at 110 °C for 24 h, then weighed for 5 g samples, transferred to porcelain bowls, and heated in the muffle furnace at 430 °C for 2 h. After annealing, the samples were transferred into a desiccator for cooling and weighed again the next day. The organic matter content was determined using mass differences. Both pH and organic matter content measurements were made of three random subsamples from the composite soil sample of each locality, and expressed as mean values.
Results
Except for one in the Mediterranean region, all localities of C. archeri were situated in the Continental NATURA 2000 biogeographical region, with no localities recorded in the Alpine region (Fig. 2). Localities were situated between 104 and 532 m. a.s.l., with 10th and 90th inter-percentile ranging from 109 to 381 m. a.s.l. (Tab. 2).
Tab. 2. Environmental variables values for Clathrus archeri localities (Tab. 1) in Croatia. pH – pH value of topsoil in 0 – 5 cm depth, elevation (m.a.s.l.), slope (°), -prec – precipitation (mm) and -temp – temperature (°C) for four seasons (sp – spring, su – summer, a – autumn, w – winter) and annual (y-). min – minimum, max – maximum, mean – average, perc10 and perc90 – 10% and 90% percentiles for each environmental variable.
The 10-90 inter-percentile range for mean annual temperature was 9.6 to 10.8 °C, and for annual precipitation 831 to 1088 mm. With respect to the seasonal precipitation amounts, there were no seasons with less than 150 mm at any locality (Tab. 2). Soil reaction ranged from 5.1 to 6.8, with 10-90 inter-percentile range being 5.3 to 6.3 (Tab. 2), both corresponding to the acidic part of the pH scale. Our measurements of soil reaction sampled at two localities in a stand of planted Eastern White Pine Pinus strobus L. (No. 3 in Tab. 1) and in the Epimedio-Carpinetum betuli (Horvat 1938) Borhidi 1963 forest (No. 24 in Tab. 1) have shown an even lower pH, of 4.3 and 4.1, respectively. Regarding organic matter content, the mean value for Donje Ladanje (No. 3 in Tab. 1) was 19.14% and for Donji Stupnik (No. 24 in Tab. 1) 16.75%, indicating that soils at both localities were rich in organic matter.
Climate-only habitat suitability model (Fig. 3) and Climate-pH habitat suitability model (Fig. 4) yielded a very similar general spatial pattern at country level, with Climate-pH based model seemingly being spatially more specific in terms of areas with high suitability.
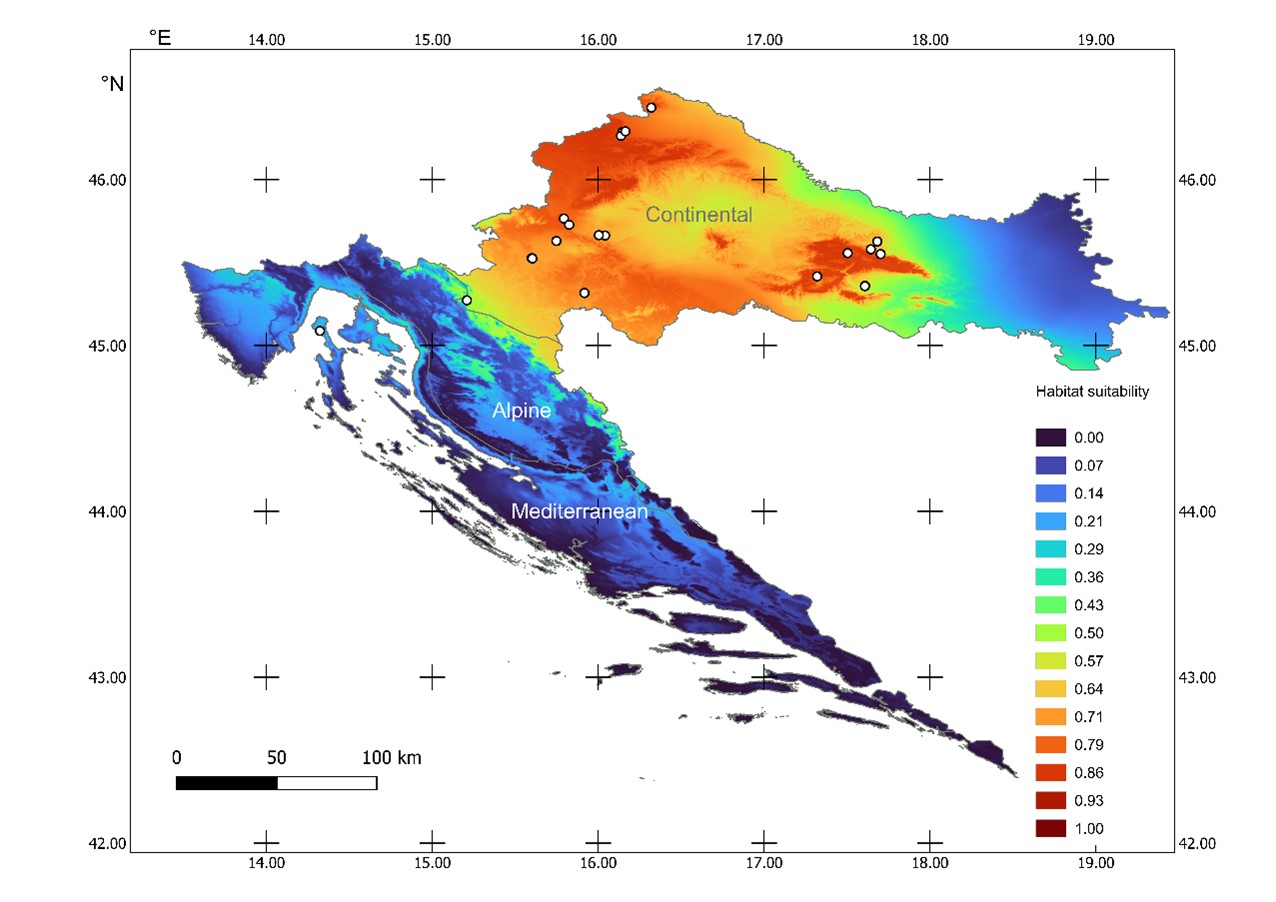
Fig. 3. Maxent habitat suitability model for Clathrus archeri in Croatia based on eight climatic variables (mean temperature and amount of precipitation per meteorological season). Colours on the map, with corresponding number indicates habitat suitability for C. archeri in range from 1 (red – presumed 100% suitable habitat) to 0 (blue – 0% suitable habitat). White circles indicate C. archeri occurrences in Croatia used to build the model, NATURA 2000 biogeographical regions indicated.
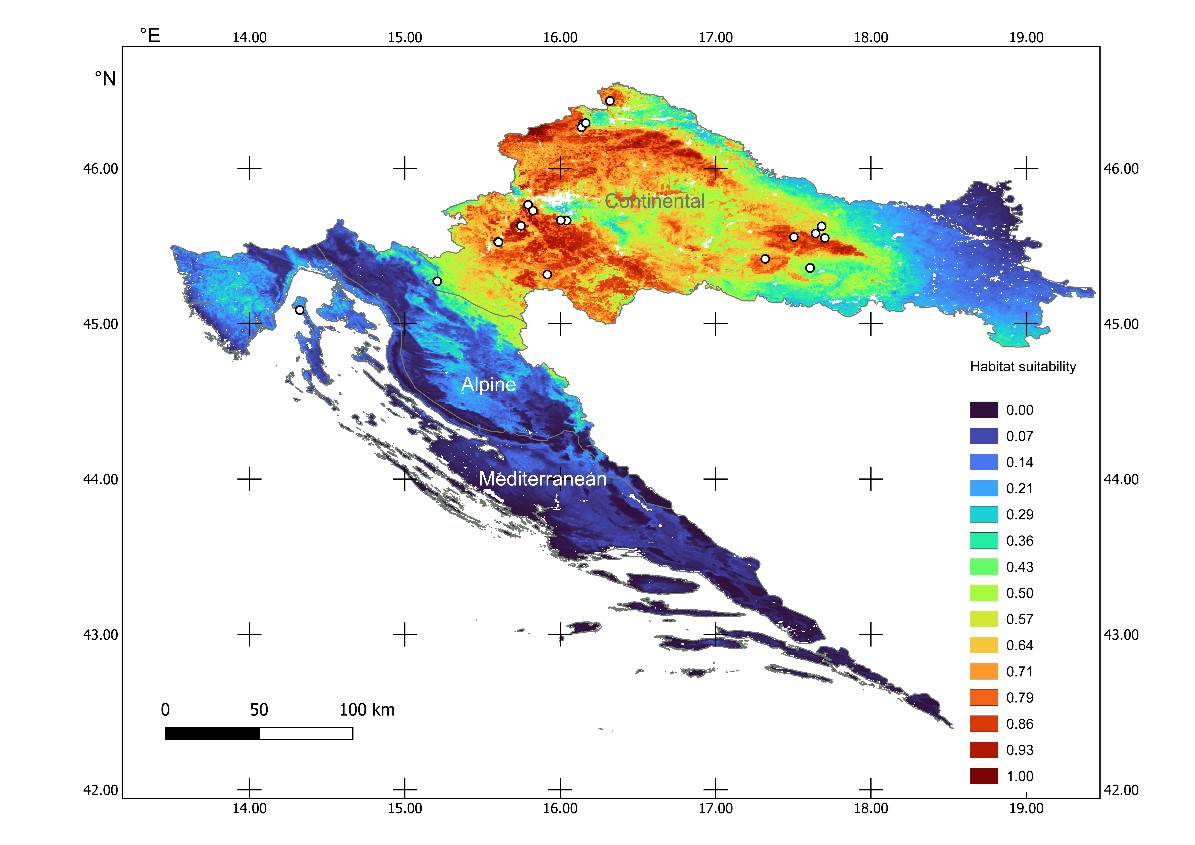
Fig. 4. Maxent habitat suitability model for Clathrus archeri in Croatia based on eight climatic variables (mean temperature and amount of precipitation per meteorological season) and pH value of soil in 0–5 cm depth. Colours on the map, with corresponding number indicates habitat suitability for C. archeri in range from 1 (red – presumed 100% suitable habitat) to 0 (blue – 0% suitable habitat). White circles indicate C. archeri occurrences in Croatia used to build the model, NATURA 2000 biogeographical regions indicated.
The latter model also contains some no-data areas, as a consequence of the lack of pH data for water body areas and human settlements. The majority of predicted suitable habitats are in the Continental biogeographical region where most of the recorded localities are situated. There are some moderately suitable areas in the Alpine region, in which, however, there are currently no records of C. archeri. Those areas are mostly in parts adjacent to the Continental region around the Kupa, Dobra and Una rivers, and in lower elevation parts corresponding to the presence of flat depressions within the karst known as “polje” (e.g. Cerničko polje, Gacko polje, Krbavsko polje). Within the Mediterranean region, very low habitat suitability was predicted by both Maxent models, with slightly higher values in the Northern Adriatic where one locality exists (No. 22 in Tab. 1). For both habitat suitability models, area under curve (AUC) of the receiver operating curve (ROC) had similar values, AUC = 0.817 for Climate-only and AUC = 0.827 for Climate-pH model (Tab. 3).
Tab. 3. Variable contributions to Maxent habitat suitability models for Clathrus archeri in Croatia. Abbreviations: w – winter, a – autumn, su – summer, sp – spring, temp – mean temperature, prec – total amount of precipitation, pH – pH value of topsoil in 0 – 5 cm depth.
Precipitation variables contributed more to the models than the temperature variables. Summer precipitation had the highest percentage of contribution to the Climate-only model, while the pH of topsoil had the highest percentage to the Climate-pH model. Winter precipitation was second best in terms of the percentage of the contribution to the models, but had by far the highest permutation importance indicating that models were heavily dependent on it (Tab. 3). On three localities that were not used for habitat suitability modelling (No. 2, 5 and 20 in Tab. 1), both models predicted high values of suitability: 0.68, 0.67 and 0.75, respectively, for Climate-only model; 0.75, 0.79 and 0.89 for Climate-pH model.
Discussion
The history and introduction of C. archeri in Croatia remain unclear. Its first mention in Božac (1984) in his popular guide to fungi cannot be considered the first evidence of its presence in Croatia because the exact geographical extent covered by the guide is not defined. At that time Croatia was a part of former Socialist Federative Republic of Yugoslavia. Furthermore, in the book’s preface the author explains that the most frequently used criterion for inclusion of fungi species in the book was their high frequency in “our regions” (which could be referred to other constituent republics of the former country beside Croatia) and in Central Europe. The latter could mean that C. archeri found its place in the book without actually being present in Croatia. However, at that time C. archeri had already been recorded (1966) in Slovenia (also part of the former country) for quite a long time (Vrščaj et al. 2004, Stanič 2024). In 1998, C. archeri was included in the first list of protected fungi in Croatia (Official Gazette 1998), and remained there in the second (published also in 1998) and third edition (published in 2002). We could not trace back the background for this inclusion, since the first mention of C. archeri in the literature is that of Tkalčec et al. (2005) in their paper on gasteral Basidiomycota of Croatia in which they note that their reference for C. archeri was the first for Croatia. Tkalčec (personal communication) found it in 1999 in two locations in the western part of Croatia. Without new evidence, this could be regarded as the year of the fungus’s first observation in Croatia. Nevertheless, given the pace of spread eastward from its initial introduction to France in 1920 (Lemasson 1923 in Parent and Thoen 1986), and subsequent first observations in Switzerland in 1942 (Vischer 1943), Austria in 1948 (Lohwag 1948) and Slovenia in 1966 (Vrščaj et al. 2004, Stanič 2024), it is possible that the fungus was present in Croatia from the 1980s or early 1990s, but remained unnoticed. Although we cannot be sure of the exact time of its arrival, with a fair amount of certainty we can consider that it arrived from the Slovenia.
The data given here show that C. archeri is much more widespread in Croatia than was considered previously, according to the scientific literature. Both habitat suitability models imply that it could occupy even larger areas than presently, at least across the majority of the Natura 2000 Continental biogeographical region in Croatia. High values of predicted habitat suitability models at locations that were not used in building the models, as well as reasonably high AUC values work in favour of the predicted models’ reliabilities. In both models, precipitation turned out to be more important than temperature in defining the suitability for C. archeri. In habitat suitability models for Romania (Bîrsan et al. 2021) and on a global level (Pietras et al. 2021) there were the same findings, with the coldest quarter of the year’s precipitation contributing the most to their models. Here, winter precipitation (which corresponds to the precipitation of the coldest quarter) was the most important among the climate variables in the Climate-pH model, while it was second most important, after summer precipitation, in the Climate-only model (Tab. 3). The Mediterranean part of Croatia has a distinct dry season during the summer, hence this could play an important role in defining potential niche of the species in Croatia. Pietras et al. (2016) found that in Poland C. archeri is mostly present in areas with annual precipitation greater than 600 mm. They confirmed its presence in part of Poland with low mean annual precipitation (550 mm), but during the year with an exceptionally wet season. The only location so far known in the Mediterranean part of Croatia is in the Northern Adriatic, which does not have such dry summer periods as those in Central and Southern parts of the Adriatic, where summer precipitation is mostly below 150 mm, which in combination with highly permeable limestone bedrock leads to dry soil conditions.
Another important aspect that could limit C. archeri in the Mediterranean part of Croatia is the soil reaction. In their review of the fungus’s presence in Europe, Parent and Thoen (1986) found it to favour acidic soils, reporting a pH range in France from 4.5 to 6, which matches our own measurements from two localities having pH value of 4.1 and 4.3, and pH values (Tab. 2) obtained from the dataset available for Europe (Poggio et al. 2021). The Croatian coast is characterised by limestone bedrock and the consequent alkaline soil reaction. In Giovanetti et al. (2014), soil reactions measured across the Croatian part of Adriatic coast, except for one sample with a pH value of 6.2, ranged from 7.7 to 8.1. The importance of the soil reaction for C. archeri is supported by the fact that it was the most important variable in the Climate-pH model. Nevertheless, given the fact that the fungus is present in the Mediterranean region, and that there is one observation (iNaturalist. org 2018) from May 2018 in the Slovenian part of the Istria peninsula very close (1.2 km) to the Croatian border, it is reasonable to expect the fungus’ presence in Istria, although neither of the models (Fig. 3 and Fig. 4) predicted high suitability there. Consequently, both models seem to underestimate the suitability in the Mediterranean region, although we believe that only the Northern part of the Adriatic, which is not characterised by as low summer precipitation and alkaline soil reaction as the more Southern areas, is concerned.
The presence of highly suitable areas in the Alpine region in which there are no known localities, does not necessarily mean that the models overestimate the suitability for C. archeri in these parts of Croatia. Indeed, there are no ecological constraints for the fungus there, since that area has a lot of precipitation as well as parts with acid soils. Although the maximum elevation of localities of C. archeri presented here is just 532 m. a.s.l., with 90% of locations appearing below 400 m. a.s.l. (Tab. 2), the species can be found at much higher elevations e.g. 1200 m. a.s.l. in Romania (Bîrsan et al. 2014), or as reported by Parent and Thoen (1986) even up to 2000 m. a.s.l. The fact that currently known localities in Croatia are mostly below 400 m. a.s.l., could be merely a consequence of prevalence of incident observations during field visits that were not targeted specifically to survey the presence of C. archeri. Some of those observations could be from edible fungi collectors who prefer this elevation belt that corresponds to distribution of pedunculate (Quercus robur L.) and sessile (Q. petraea (Matt.) Liebl.) oak forests, known for their fungi richness. An additional explanation could be the very short duration of the fungus’ visible presence which lasts approximately just one week per individual sporocarp. This includes the egg phase, which can be observed up to 5 days. However, without dissection it is impossible to record an observation, since other stinkhorn fungi also develop eggs. We expect that sampling effort (in this case actually simply the number of field days, given the non-targeted nature of it) is much higher in lower elevations, hence the higher probability of observing C. archeri there.
In the future, we could expect an increase in the presence of C. archeri in Croatia in terms of its abundance and occupancy area. Even at present, on various social networks photographs of the fungus from Croatia can be found, but without details on its geographical origin, hence they could not be used here. It is not easy to predict how much this increase might be, and how long it would last. In their analysis of the impact of climate change on C. archeri, Pietras et al. (2021) used three scenarios for the year 2080 expecting the fungus’ range contraction in Central Europe, including the Continental region of Croatia from which currently the majority of observations derive. Depending on the climate change scenario employed, there could be some range expansion in the areas that overlap with the Mediterranean region in Croatia. However, Pietras et al. (2021) developed their models exclusively with bioclimatic variables, without data on soil acidity, which could play a significant role in shaping the future of C. archeri distribution.
Almost 20 years after its first mention for Croatia in the scientific literature (Tkalčec et al. 2005) and just one short note in the meantime reporting on two localities (Kranjčev 2008) we reveal that C. archeri is nowadays, in terms of its spatial distribution, a widespread alien fungus in Croatia. Open access data provided by international networks like GBIF and iNaturalist were indispensable for this analysis. Consequently, these data should be an integral part in further efforts in increasing our knowledge on alien fungi, and other taxa, presence and distribution in Croatia, that are otherwise poorly covered in the scientific literature. In addition, efforts to raise the awareness of such species among colleagues and the general public, as well as of the importance of observations being deposited on platforms like GBIF and/or iNaturalist could significantly contribute to the amount of available observational records. Habitat suitability models can help in planning field surveys, which is especially useful for taxa with low cost-efficiency field work, as C. archeri is, due to its short sporocarp longevity.
Acknowledgements
This paper was written in support of the journal Acta Botanica Croatica on the occasion of its 100th anniversary.
We would like to thank iNaturalist observers Nevio Biškupić, Marko Doboš, Eva Janeković, Sara Srša and Edi Thür for granting permission to use their data in this paper. The editor and two reviewers have provided constructive and valuable comments that helped us improve the manuscript, for which we are grateful. This research was funded by the University of Zagreb, Croatia, grant number 20286538–Distribution and ecological profiles of invasive plants.